

Role for stromal heterogeneity in prostate tumorigenesis
- PMID: 21444670
- PMCID: PMC3096737
- DOI: 10.1158/0008-5472.CAN-10-2999
Role for stromal heterogeneity in prostate tumorigenesis
Abstract
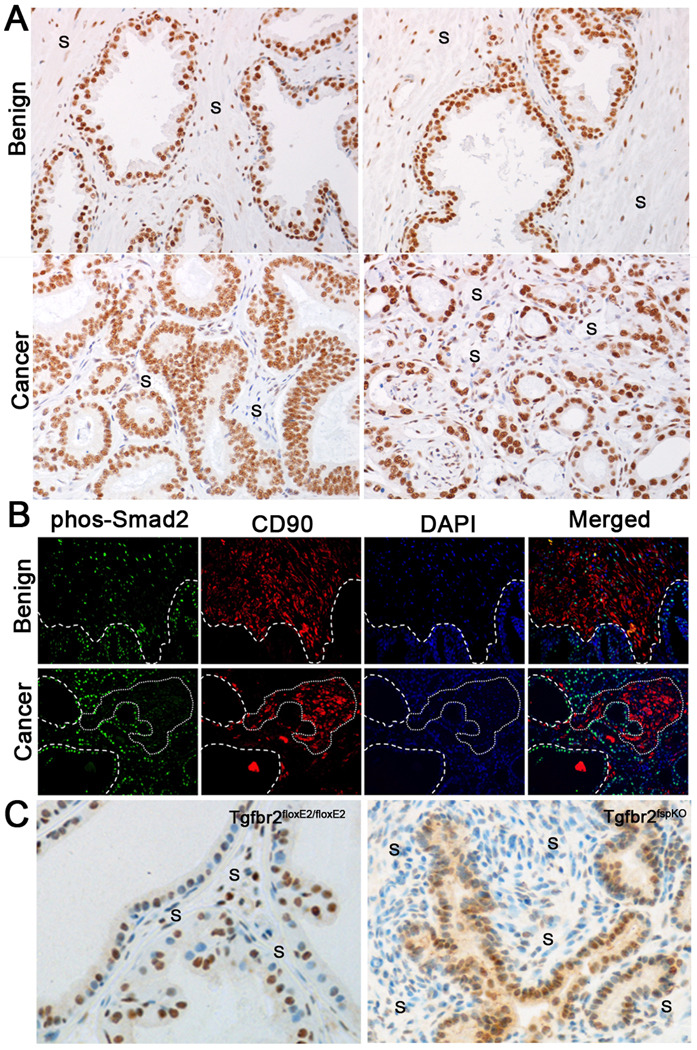
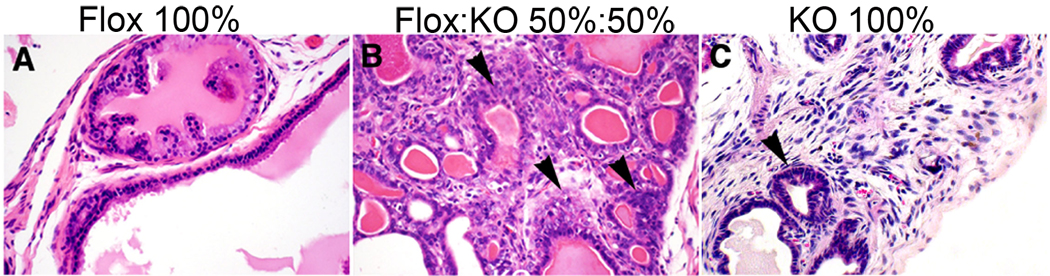
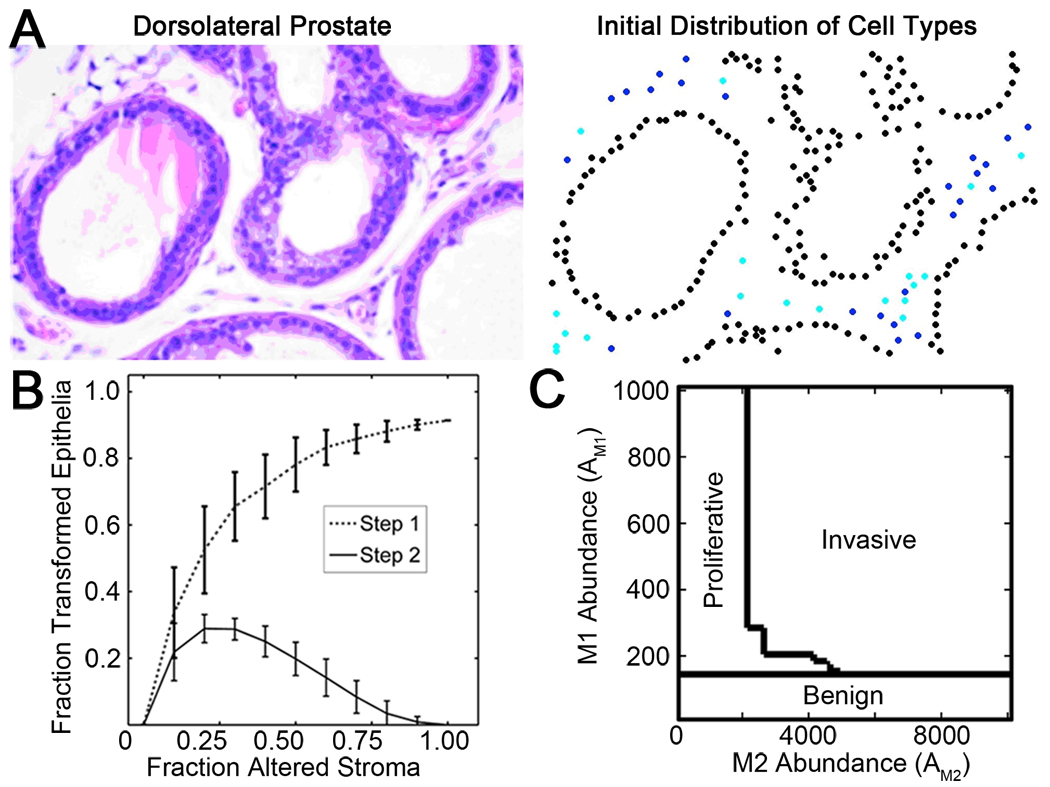
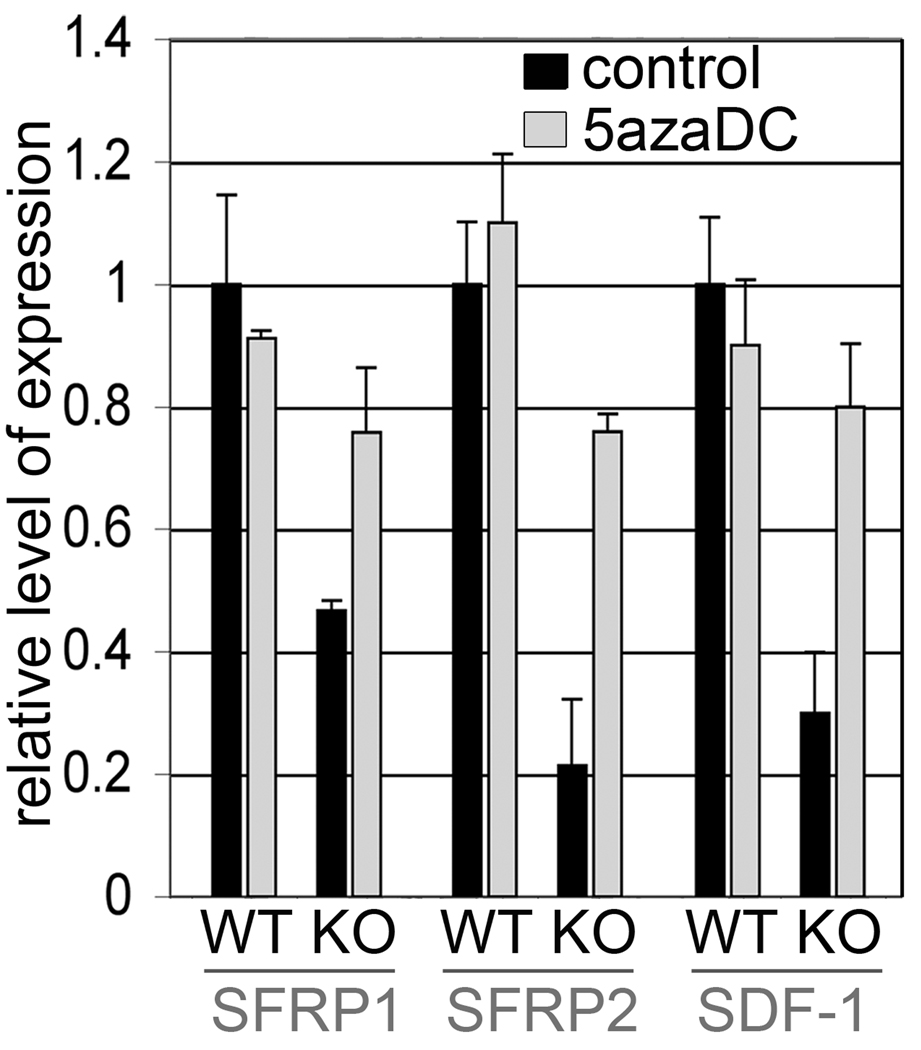
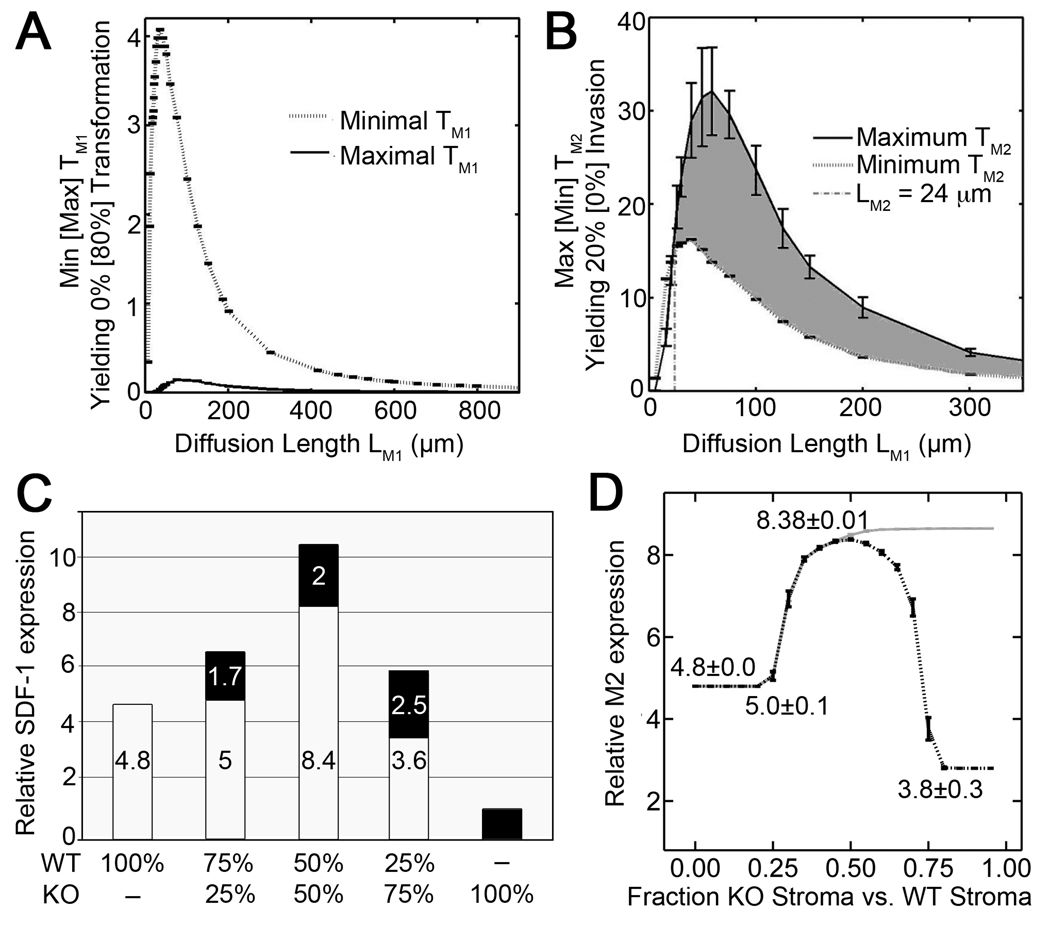
Prostate cancer develops through a stochastic mechanism whereby precancerous lesions on occasion progress to multifocal adenocarcinoma. Analysis of human benign and cancer prostate tissues revealed heterogeneous loss of TGF-β signaling in the cancer-associated stromal fibroblastic cell compartment. To test the hypothesis that prostate cancer progression is dependent on the heterogeneous TGF-β responsive microenvironment, a tissue recombination experiment was designed in which the ratio of TGF-β responsive and nonresponsive stromal cells was varied. Although 100% TGF-β responsive stromal cells supported benign prostate growth and 100% TGF-β nonresponsive stromal cells resulted in precancerous lesions, only the mixture of TGF-β responsive and nonresponsive stromal cells resulted in adenocarcinoma. A computational model was used to resolve a mechanism of tumorigenic progression in which proliferation and invasion occur in two independent steps mediated by distinct stromally derived paracrine signals produced by TGF-β nonresponsive and responsive stromal cells. Complex spatial relationships of stromal and epithelial cells were incorporated into the model on the basis of experimental data. Informed by incorporation of experimentally derived spatial parameters for complex stromal-epithelial relationships, the computational model indicated ranges for the relative production of paracrine factors by each cell type and provided bounds for the diffusive range of the molecules. Because SDF-1 satisfied model predictions for an invasion-promoting paracrine factor, a more focused computational model was subsequently used to investigate whether SDF-1 was the invasion signal. Simulations replicating SDF-1 expression data revealed the requirement for cooperative SDF-1 expression, a prediction supported biologically by heterotypic stromal interleukin-1β signaling between fibroblastic cell populations. The cancer stromal field effect supports a functional role for the unaltered fibroblasts as a cooperative mediator of cancer progression.
©2011 AACR
Figures

Similar articles
-
Cross-talk between paracrine-acting cytokine and chemokine pathways promotes malignancy in benign human prostatic epithelium.Cancer Res. 2007 May 1;67(9):4244-53. doi: 10.1158/0008-5472.CAN-06-3946. Cancer Res. 2007. PMID: 17483336
-
Prostate tumor progression is mediated by a paracrine TGF-beta/Wnt3a signaling axis.Oncogene. 2008 Nov 27;27(56):7118-30. doi: 10.1038/onc.2008.293. Epub 2008 Aug 25. Oncogene. 2008. PMID: 18724388 Free PMC article.
-
Altered TGF-β signaling in a subpopulation of human stromal cells promotes prostatic carcinogenesis.Cancer Res. 2011 Feb 15;71(4):1272-81. doi: 10.1158/0008-5472.CAN-10-3142. Epub 2011 Feb 8. Cancer Res. 2011. PMID: 21303979 Free PMC article.
-
Transforming growth factor-beta in benign and malignant prostate.Prostate. 1999 Jun 1;39(4):285-90. doi: 10.1002/(sici)1097-0045(19990601)39:4<285::aid-pros9>3.0.co;2-7. Prostate. 1999. PMID: 10344218 Review.
-
Peptide growth factors in the prostate as mediators of stromal epithelial interaction.Br J Urol. 1996 May;77(5):627-33. doi: 10.1046/j.1464-410x.1996.09721.x. Br J Urol. 1996. PMID: 8689101 Review.
Cited by
-
Sonic hedgehog signals to multiple prostate stromal stem cells that replenish distinct stromal subtypes during regeneration.Proc Natl Acad Sci U S A. 2013 Dec 17;110(51):20611-6. doi: 10.1073/pnas.1315729110. Epub 2013 Nov 11. Proc Natl Acad Sci U S A. 2013. PMID: 24218555 Free PMC article.
-
TGFβ loss activates ADAMTS-1-mediated EGF-dependent invasion in a model of esophageal cell invasion.Exp Cell Res. 2015 Jan 1;330(1):29-42. doi: 10.1016/j.yexcr.2014.07.021. Epub 2014 Jul 24. Exp Cell Res. 2015. PMID: 25064463 Free PMC article.
-
Cancer: a mirrored room between tumor bulk and tumor microenvironment.J Exp Clin Cancer Res. 2021 Jun 28;40(1):217. doi: 10.1186/s13046-021-02022-5. J Exp Clin Cancer Res. 2021. PMID: 34183054 Free PMC article. Review.
-
Cancer-Associated Fibroblast Heterogeneity, Activation and Function: Implications for Prostate Cancer.Biomolecules. 2022 Dec 29;13(1):67. doi: 10.3390/biom13010067. Biomolecules. 2022. PMID: 36671452 Free PMC article. Review.
-
The reactive stroma microenvironment and prostate cancer progression.Endocr Relat Cancer. 2012 Oct 30;19(6):R187-204. doi: 10.1530/ERC-12-0085. Print 2012 Dec. Endocr Relat Cancer. 2012. PMID: 22930558 Free PMC article. Review.
References
-
- Thesleff I, Vaahtokari A, Partanen AM. Regulation of organogenesis. Common molecular mechanisms regulating the development of teeth and other organs. Int J Dev Biol. 1995;39:35–50. - PubMed
-
- Bostwick DG, Pacelli A, Lopez-Beltran A. Molecular biology of prostatic intraepithelial neoplasia. Prostate. 1996;29:117–134. - PubMed
-
- Chung LW, Baseman A, Assikis V, Zhau HE. Molecular insights into prostate cancer progression: the missing link of tumor microenvironment. J Urol. 2005;173:10–20. - PubMed
-
- Alberti C. Genetic and microenvironmental implications in prostate cancer progression and metastasis. Eur Rev Med Pharmacol Sci. 2008;12:167–175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
