

Tumor-evoked regulatory B cells promote breast cancer metastasis by converting resting CD4⁺ T cells to T-regulatory cells
- PMID: 21444674
- PMCID: PMC3096701
- DOI: 10.1158/0008-5472.CAN-10-4316
Tumor-evoked regulatory B cells promote breast cancer metastasis by converting resting CD4⁺ T cells to T-regulatory cells
Abstract
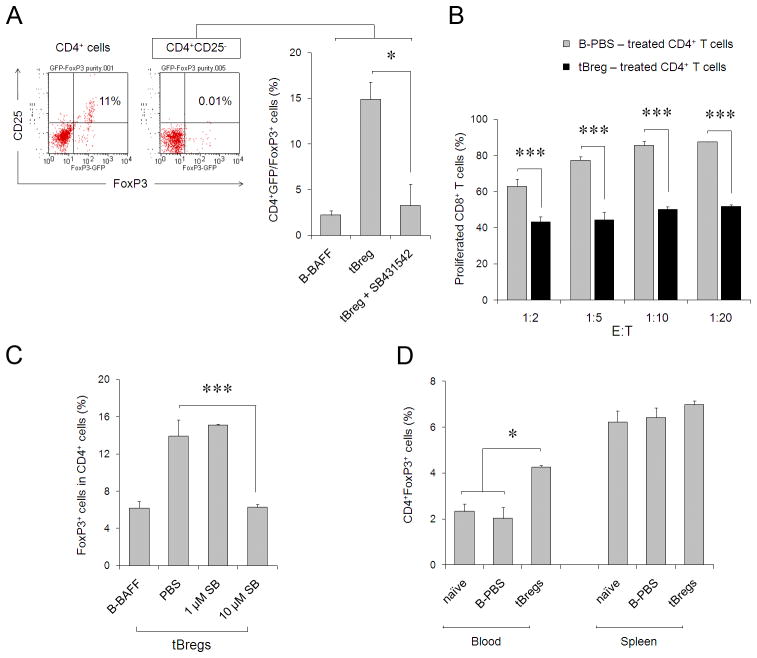
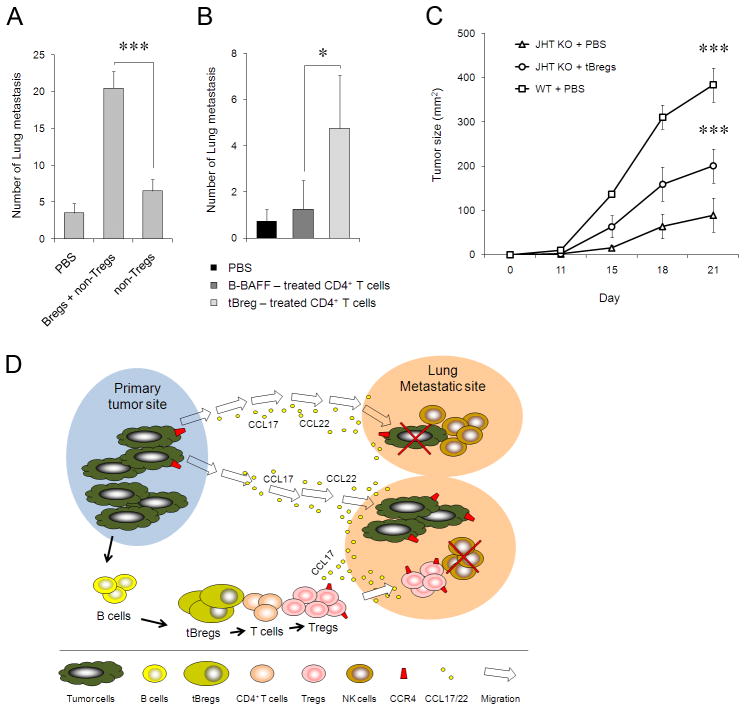
Pulmonary metastasis of breast cancer requires recruitment and expansion of T-regulatory cells (Treg) that promote escape from host protective immune cells. However, it remains unclear precisely how tumors recruit Tregs to support metastatic growth. Here we report the mechanistic involvement of a unique and previously undescribed subset of regulatory B cells. These cells, designated tumor-evoked Bregs (tBreg), phenotypically resemble activated but poorly proliferative mature B2 cells (CD19(+) CD25(High) CD69(High)) that express constitutively active Stat3 and B7-H1(High) CD81(High) CD86(High) CD62L(Low) IgM(Int). Our studies with the mouse 4T1 model of breast cancer indicate that the primary role of tBregs in lung metastases is to induce TGF-β-dependent conversion of FoxP3(+) Tregs from resting CD4(+) T cells. In the absence of tBregs, 4T1 tumors cannot metastasize into the lungs efficiently due to poor Treg conversion. Our findings have important clinical implications, as they suggest that tBregs must be controlled to interrupt the initiation of a key cancer-induced immunosuppressive event that is critical to support cancer metastasis.
©2011 AACR
Conflict of interest statement
Figures
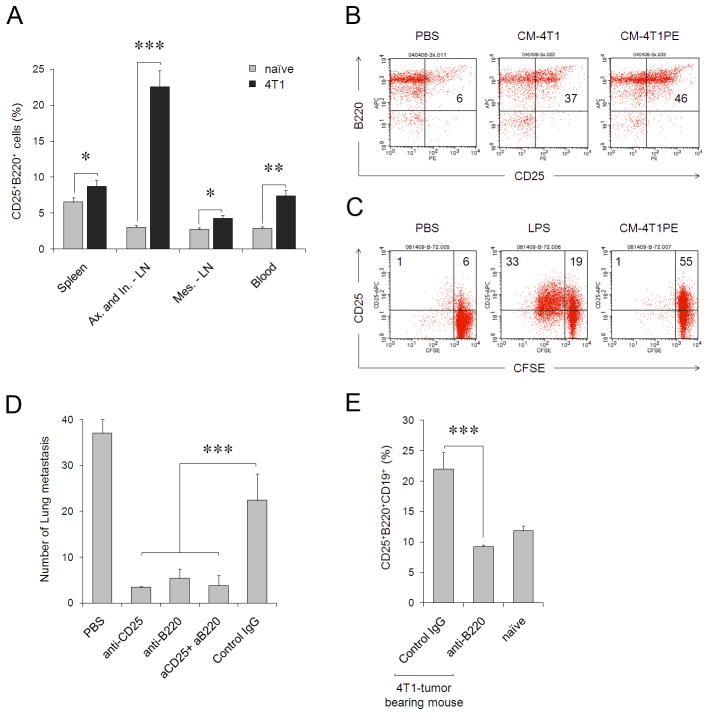
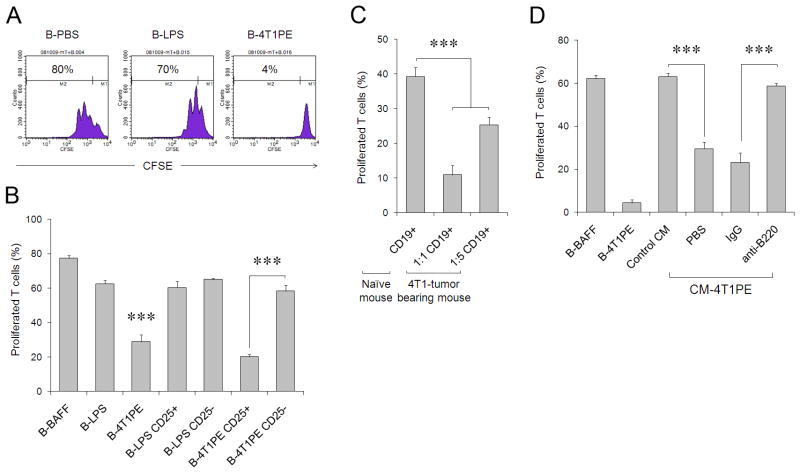
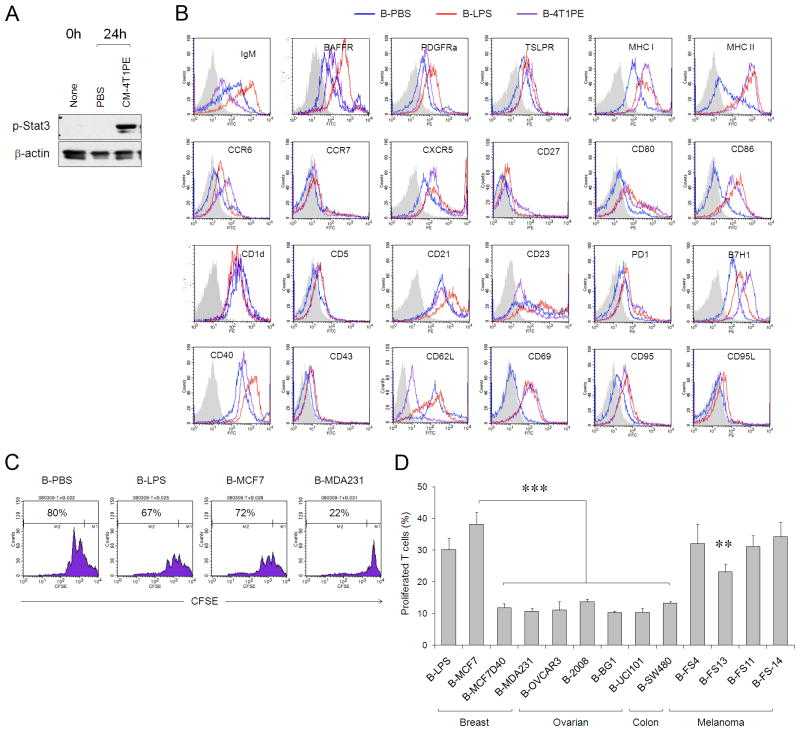
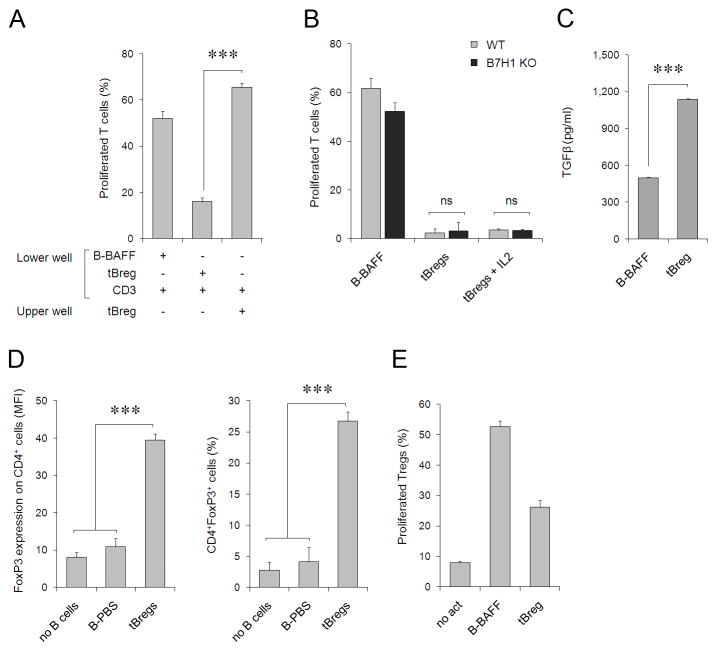
References
-
- Liu VC, Wong LY, Jang T, Shah AH, Park I, Yang X, et al. Tumor evasion of the immune system by converting CD4+ J Immunol. 2007;178:2883–92. - PubMed
-
- Lelekakis M, Moseley JM, Martin TJ, Hards D, Williams E, Ho P, et al. A novel orthotopic model of breast cancer metastasis to bone. Clin Exp Metastasis. 1999;17:163–70. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
