

Distinct mechanisms of ferritin delivery to lysosomes in iron-depleted and iron-replete cells
- PMID: 21444722
- PMCID: PMC3133360
- DOI: 10.1128/MCB.01437-10
Distinct mechanisms of ferritin delivery to lysosomes in iron-depleted and iron-replete cells
Abstract
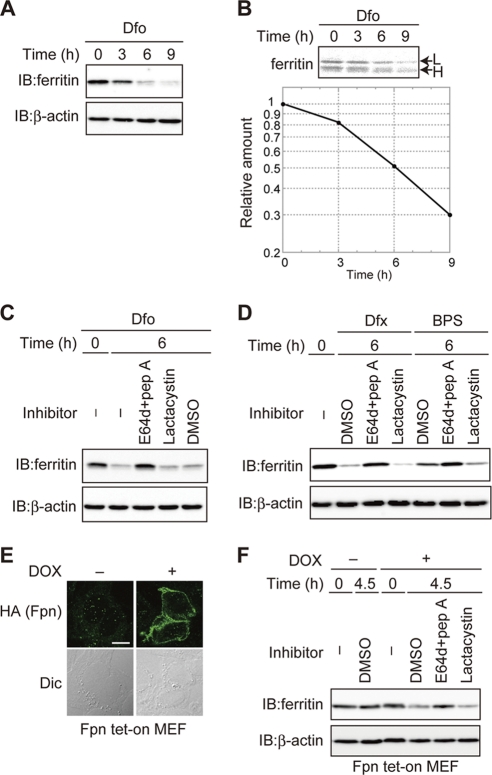
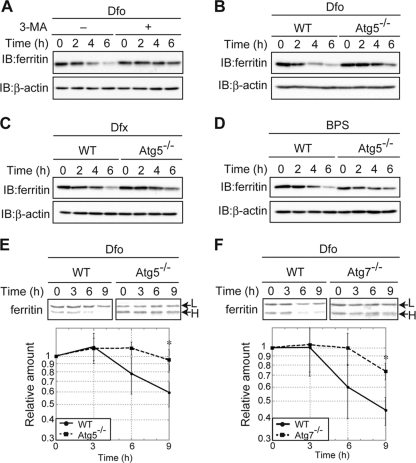
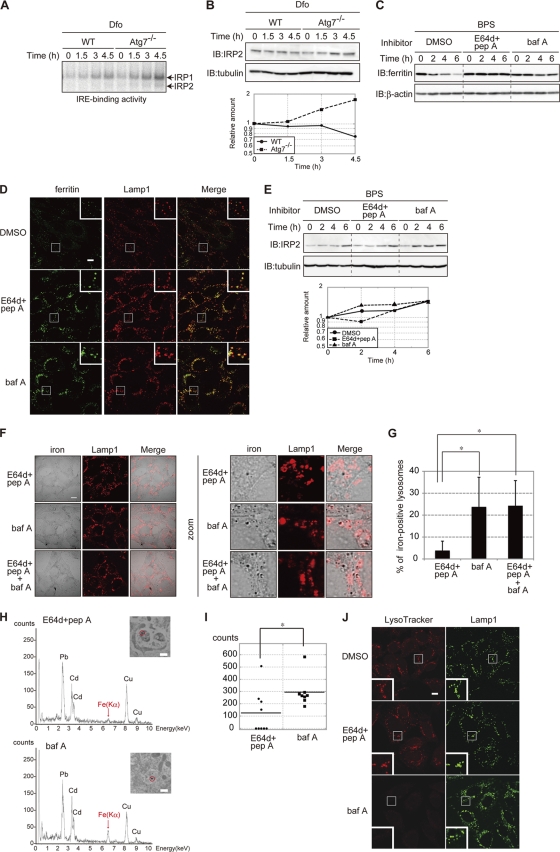
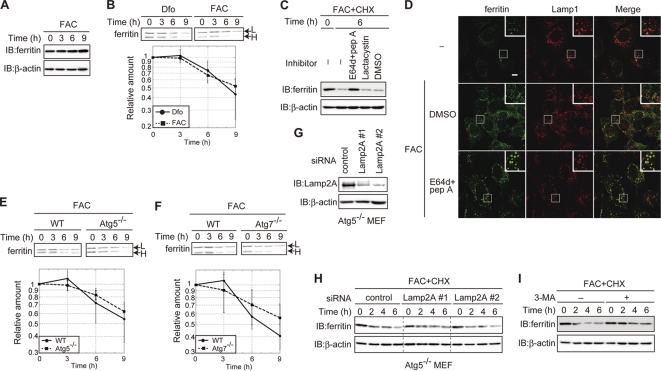
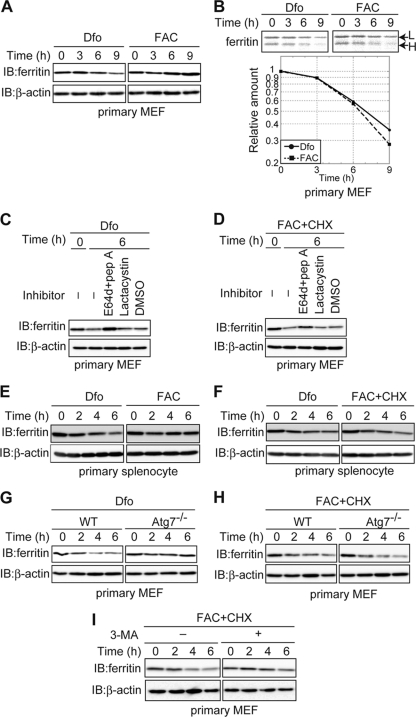
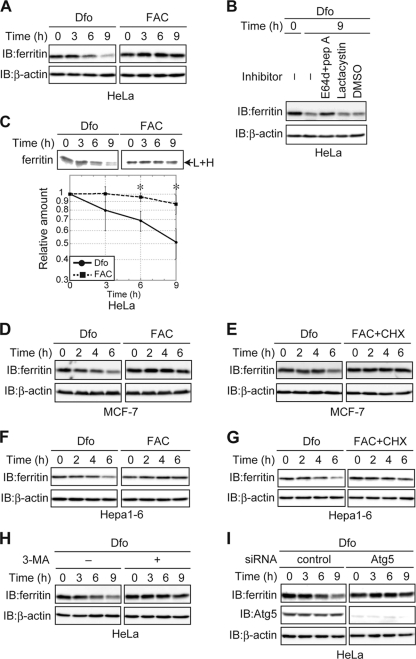
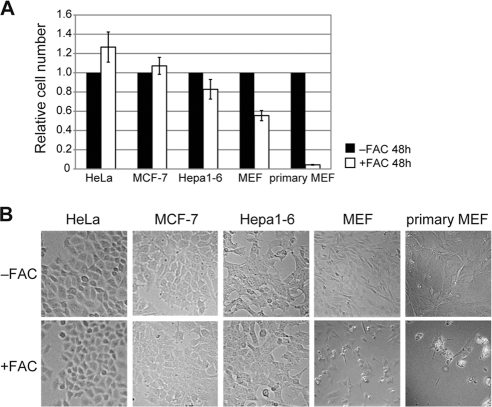
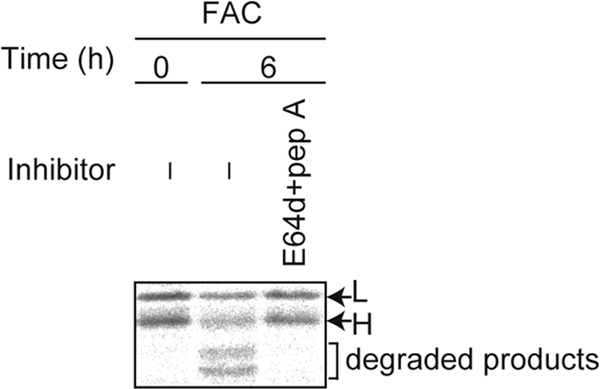
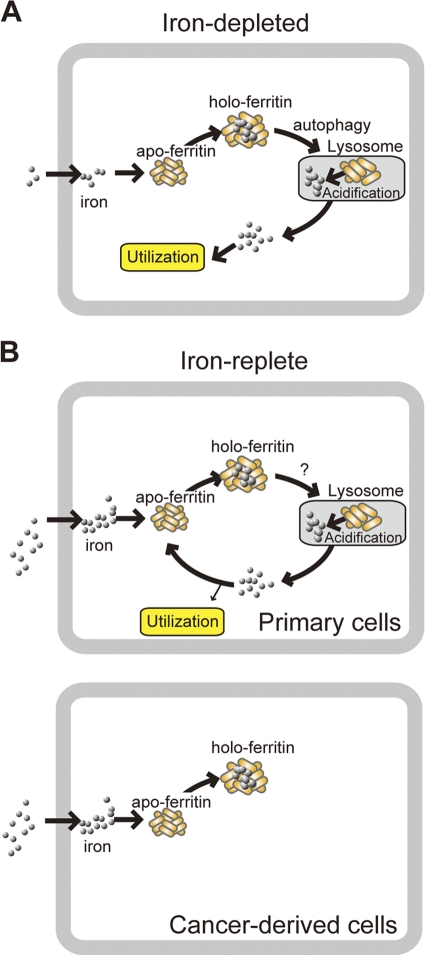
Ferritin is a cytosolic protein that stores excess iron, thereby protecting cells from iron toxicity. Ferritin-stored iron is believed to be utilized when cells become iron deficient; however, the mechanisms underlying the extraction of iron from ferritin have yet to be fully elucidated. Here, we demonstrate that ferritin is degraded in the lysosome under iron-depleted conditions and that the acidic environment of the lysosome is crucial for iron extraction from ferritin and utilization by cells. Ferritin was targeted for degradation in the lysosome even under iron-replete conditions in primary cells; however, the mechanisms underlying lysosomal targeting of ferritin were distinct under depleted and replete conditions. In iron-depleted cells, ferritin was targeted to the lysosome via a mechanism that involved autophagy. In contrast, lysosomal targeting of ferritin in iron-replete cells did not involve autophagy. The autophagy-independent pathway of ferritin delivery to lysosomes was deficient in several cancer-derived cells, and cancer-derived cell lines are more resistant to iron toxicity than primary cells. Collectively, these results suggest that ferritin trafficking may be differentially regulated by cell type and that loss of ferritin delivery to the lysosome under iron-replete conditions may be related to oncogenic cellular transformation.
Figures
References
-
- Ahlberg J., Glaumann H. 1985. Uptake microautophagy and degradation of exogenous proteins by isolated rat-liver lysosomes—effects of pH, ATP, and inhibitors of proteolysis. Exp. Mol. Pathol. 42:78–88 - PubMed
-
- Andrews N. C. 2005. Molecular control of iron metabolism. Best Pract. Res. Clin. Haematol. 18:159–169 - PubMed
-
- Arosio P., Ingrassia R., Cavadini P. 2009. Ferritins: a family of molecules for iron storage, antioxidation and more. Biochim. Biophys. Acta 1790:589–599 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical