

Mis17 is a regulatory module of the Mis6-Mal2-Sim4 centromere complex that is required for the recruitment of CenH3/CENP-A in fission yeast
- PMID: 21445296
- PMCID: PMC3061866
- DOI: 10.1371/journal.pone.0017761
Mis17 is a regulatory module of the Mis6-Mal2-Sim4 centromere complex that is required for the recruitment of CenH3/CENP-A in fission yeast
Abstract
Background: The centromere is the chromosome domain on which the mitotic kinetochore forms for proper segregation. Deposition of the centromeric histone H3 (CenH3, CENP-A) is vital for the formation of centromere-specific chromatin. The Mis6-Mal2-Sim4 complex of the fission yeast S. pombe is required for the recruitment of CenH3 (Cnp1), but its function remains obscure.
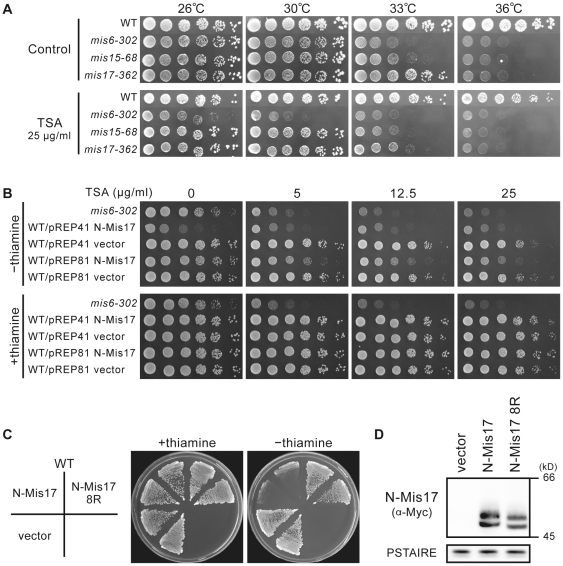
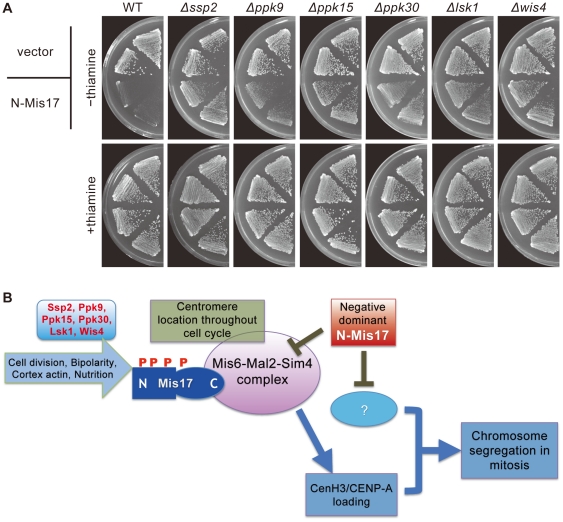
Methodology/principal findings: Mass spectrometry was performed on the proteins precipitated with Mis6- and Mis17-FLAG. The results together with the previously identified Sim4- and Mal2-TAP precipitated proteins indicated that the complex contains 12 subunits, Mis6, Sim4, Mal2, Mis15, Mis17, Cnl2, Fta1-4, Fta6-7, nine of which have human centromeric protein (CENP) counterparts. Domain dissection indicated that the carboxy-half of Mis17 is functional, while its amino-half is regulatory. Overproduction of the amino-half caused strong negative dominance, which led to massive chromosome missegregation and hypersensitivity to the histone deacetylase inhibitor TSA. Mis17 was hyperphosphorylated and overproduction-induced negative dominance was abolished in six kinase-deletion mutants, ssp2 (AMPK), ppk9 (AMPK), ppk15 (Yak1), ppk30 (Ark1), wis4 (Ssk2), and lsk1 (P-TEFb).
Conclusions: Mis17 may be a regulatory module of the Mis6 complex. Negative dominance of the Mis17 fragment is exerted while the complex and CenH3 remain at the centromere, a result that differs from the mislocalization seen in the mis17-362 mutant. The known functions of the kinases suggest an unexpected link between Mis17 and control of the cortex actin, nutrition, and signal/transcription. Possible interpretations are discussed.
Conflict of interest statement
Figures
Similar articles
-
Negative Regulation of the Mis17-Mis6 Centromere Complex by mRNA Decay Pathway and EKC/KEOPS Complex in Schizosaccharomyces pombe.G3 (Bethesda). 2019 Jun 5;9(6):1815-1823. doi: 10.1534/g3.119.400227. G3 (Bethesda). 2019. PMID: 30967422 Free PMC article.
-
Kinetochore Components Required for Centromeric Chromatin Assembly Are Impacted by Msc1 in Schizosaccharomyces pombe.Genetics. 2017 Oct;207(2):559-569. doi: 10.1534/genetics.117.300183. Epub 2017 Aug 21. Genetics. 2017. PMID: 28827290 Free PMC article.
-
Eic1 links Mis18 with the CCAN/Mis6/Ctf19 complex to promote CENP-A assembly.Open Biol. 2014 Apr 30;4(4):140043. doi: 10.1098/rsob.140043. Open Biol. 2014. PMID: 24789708 Free PMC article.
-
Two distinct pathways responsible for the loading of CENP-A to centromeres in the fission yeast cell cycle.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):595-606; discussion 606-7. doi: 10.1098/rstb.2004.1614. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897182 Free PMC article. Review.
-
Does a GATA factor make the bed for centromeric nucleosomes?Cell Cycle. 2003 Jul-Aug;2(4):277-8. Cell Cycle. 2003. PMID: 12851470 Review.
Cited by
-
Molecular basis for inner kinetochore configuration through RWD domain-peptide interactions.EMBO J. 2017 Dec 1;36(23):3458-3482. doi: 10.15252/embj.201796636. Epub 2017 Oct 18. EMBO J. 2017. PMID: 29046335 Free PMC article.
-
Mutation of histone H3 serine 86 disrupts GATA factor Ams2 expression and precise chromosome segregation in fission yeast.Sci Rep. 2015 Sep 15;5:14064. doi: 10.1038/srep14064. Sci Rep. 2015. PMID: 26369364 Free PMC article.
-
The ABCs of CENPs.Chromosoma. 2011 Oct;120(5):425-46. doi: 10.1007/s00412-011-0330-0. Epub 2011 Jul 13. Chromosoma. 2011. PMID: 21751032 Review.
-
Set2 regulates Ccp1 and Swc2 to ensure centromeric stability by retargeting CENP-A.Nucleic Acids Res. 2024 May 8;52(8):4198-4214. doi: 10.1093/nar/gkae084. Nucleic Acids Res. 2024. PMID: 38442274 Free PMC article.
-
Inner Kinetochore Protein Interactions with Regional Centromeres of Fission Yeast.Genetics. 2015 Oct;201(2):543-61. doi: 10.1534/genetics.115.179788. Epub 2015 Aug 13. Genetics. 2015. PMID: 26275423 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
