

DUX4, a candidate gene for facioscapulohumeral muscular dystrophy, causes p53-dependent myopathy in vivo
- PMID: 21446026
- PMCID: PMC4098764
- DOI: 10.1002/ana.22275
DUX4, a candidate gene for facioscapulohumeral muscular dystrophy, causes p53-dependent myopathy in vivo
Abstract
Objective: Facioscapulohumeral muscular dystrophy (FSHD) is associated with D4Z4 repeat contraction on human chromosome 4q35. This genetic lesion does not result in complete loss or mutation of any gene. Consequently, the pathogenic mechanisms underlying FSHD have been difficult to discern. In leading FSHD pathogenesis models, D4Z4 contractions are proposed to cause epigenetic changes, which ultimately increase expression of genes with myopathic potential. Although no gene has been conclusively linked to FSHD development, recent evidence supports a role for the D4Z4-encoded DUX4 gene in FSHD. In this study, our objective was to test the in vivo myopathic potential of DUX4.
Methods: We delivered DUX4 to zebrafish and mouse muscle by transposon-mediated transgenesis and adeno-associated viral vectors, respectively.
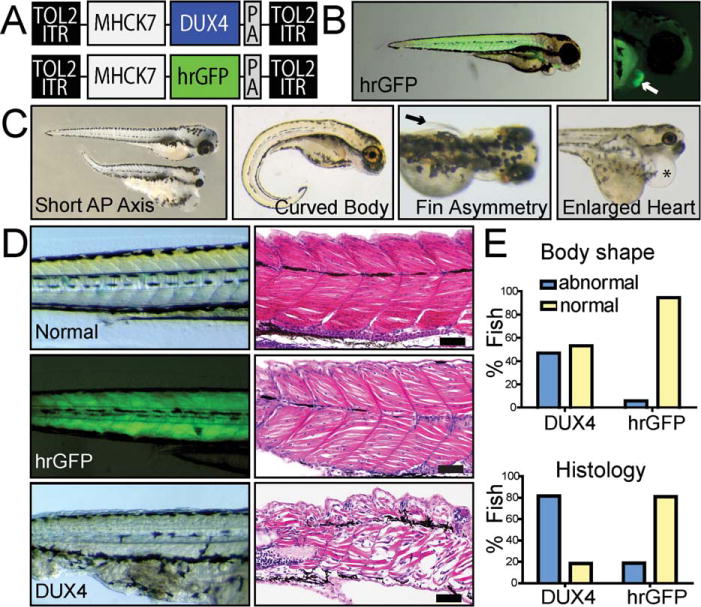
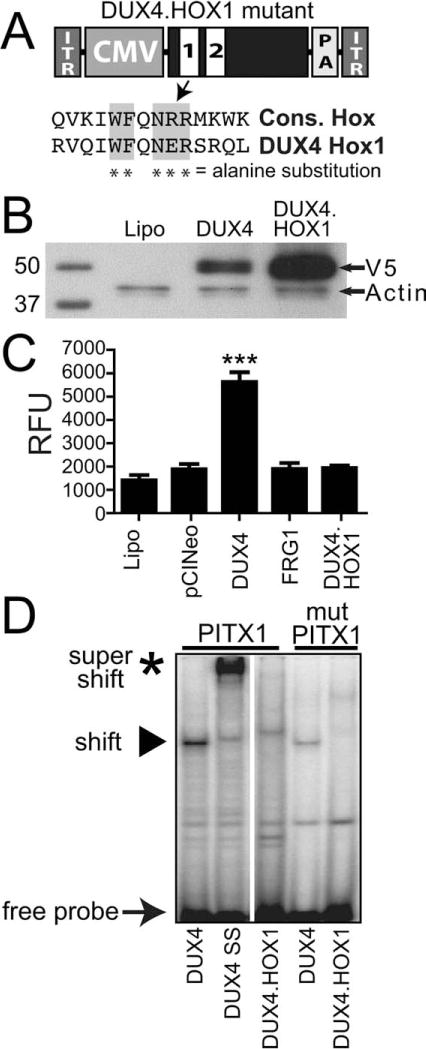
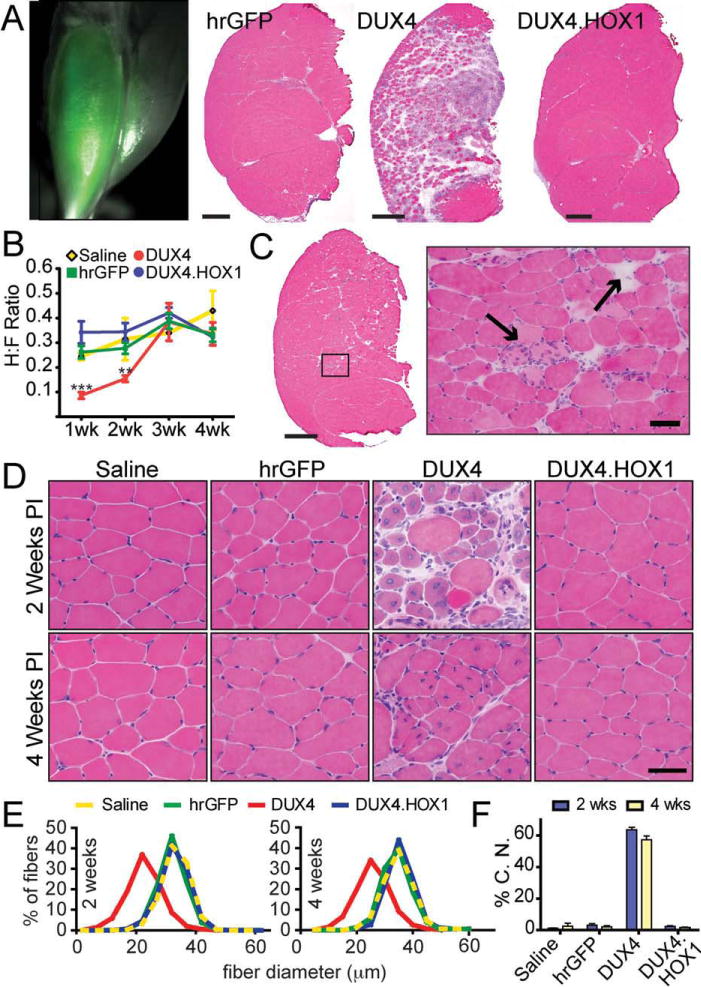
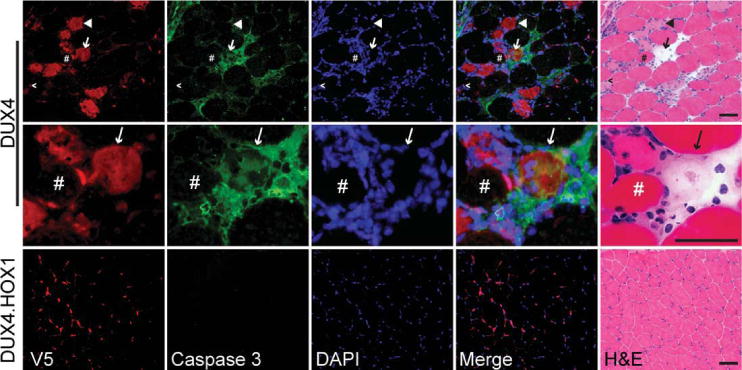
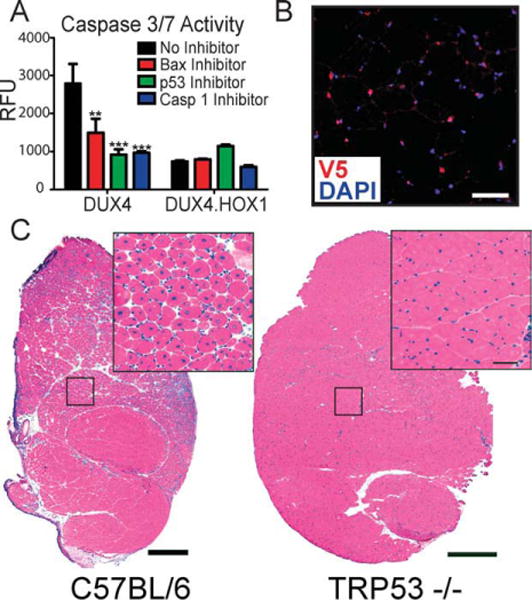
Results: Overexpression of DUX4, which encodes a transcription factor, caused abnormalities associated with muscular dystrophy in zebrafish and mice. This toxicity required DNA binding, because a DUX4 DNA binding domain mutant produced no abnormalities. Importantly, we found the myopathic effects of DUX4 were p53 dependent, as p53 inhibition mitigated DUX4 toxicity in vitro, and muscles from p53 null mice were resistant to DUX4-induced damage.
Interpretation: Our work demonstrates the myopathic potential of DUX4 in animal muscle. Considering previous studies showed DUX4 was elevated in FSHD patient muscles, our data support the hypothesis that DUX4 overexpression contributes to FSHD development. Moreover, we provide a p53-dependent mechanism for DUX4 toxicity that is consistent with previous studies showing p53 pathway activation in FSHD muscles. Our work justifies further investigation of DUX4 and the p53 pathway in FSHD pathogenesis.
Copyright © 2010 American Neurological Association.
Conflict of interest statement
J.Y.: grants/grants pending, NIH. S.Q.H.: grants/grants pending, NIH.
Figures
References
-
- Flanigan KM. Myology. 3. New York, NY: McGraw-Hill Medical Publishing Division; 2004. Facioscapulohumeral muscular dystrophy and scapuloperoneal disorders; pp. 1123–1133.
-
- Prevalence of rare diseases: bibliographic data. Orphanet. 2008 Available at: http://www.orpha.net/orphacom/cahiers/docs/GB/Prevalence_of_rare_disease... Accessed May 2010.
-
- Klinge L, Eagle M, Haggerty ID, et al. Severe phenotype in infantile facioscapulohumeral muscular dystrophy. Neuromuscul Disord. 2006;16:553–558. - PubMed
-
- van Deutekom JC, Wijmenga C, van Tienhoven EA, et al. FSHD associated DNA rearrangements are due to deletions of integral copies of a 3.2 kb tandem repeated unit. Hum Mol Genet. 1993;2:2037–2042. - PubMed
-
- Wijmenga C, Hewitt JE, Sandkuijl LA, et al. Chromosome 4q DNA rearrangements associated with facioscapulohumeral muscular dystrophy. Nat Genet. 1992;2:26–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
