

Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems
- PMID: 21447792
- PMCID: PMC3082258
- DOI: 10.1105/tpc.110.082792
Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems
Abstract
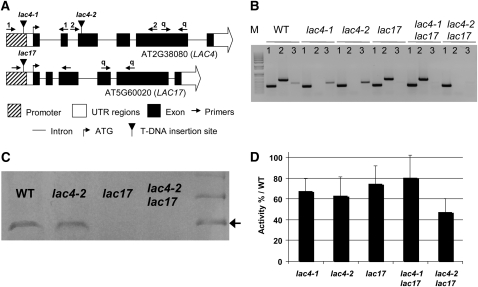
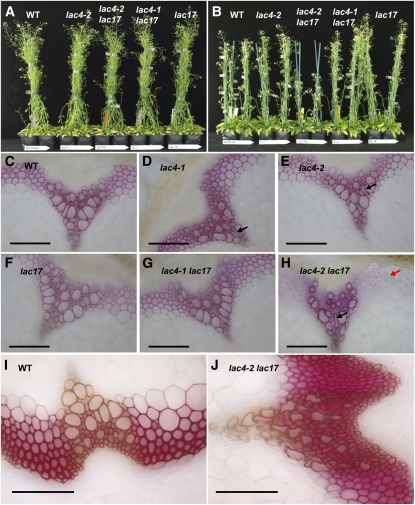
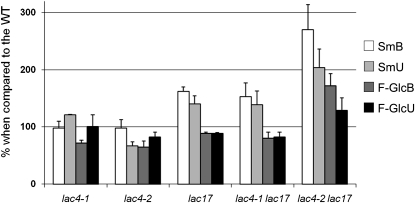
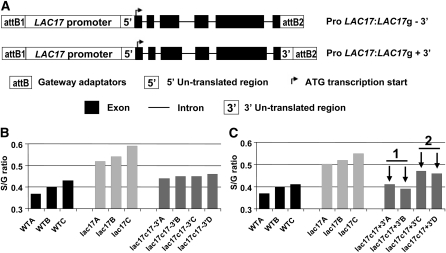
Peroxidases have been shown to be involved in the polymerization of lignin precursors, but it remains unclear whether laccases (EC 1.10.3.2) participate in constitutive lignification. We addressed this issue by studying laccase T-DNA insertion mutants in Arabidopsis thaliana. We identified two genes, LAC4 and LAC17, which are strongly expressed in stems. LAC17 was mainly expressed in the interfascicular fibers, whereas LAC4 was expressed in vascular bundles and interfascicular fibers. We produced two double mutants by crossing the LAC17 (lac17) mutant with two LAC4 mutants (lac4-1 and lac4-2). The single and double mutants grew normally in greenhouse conditions. The single mutants had moderately low lignin levels, whereas the stems of lac4-1 lac17 and lac4-2 lac17 mutants had lignin contents that were 20 and 40% lower than those of the control, respectively. These lower lignin levels resulted in higher saccharification yields. Thioacidolysis revealed that disrupting LAC17 principally affected the deposition of G lignin units in the interfascicular fibers and that complementation of lac17 with LAC17 restored a normal lignin profile. This study provides evidence that both LAC4 and LAC17 contribute to the constitutive lignification of Arabidopsis stems and that LAC17 is involved in the deposition of G lignin units in fibers.
Figures


References
-
- Adler E. (1977). Lignin chemistry. Past, present and future. Wood Sci. Technol. 11: 169–218
-
- Bao W., O’malley D.M., Whetten R., Sederoff R.R. (1993). A laccase associated with lignification in loblolly pine xylem. Science 260: 672–674 - PubMed
-
- Baudry A., Caboche M., Lepiniec L. (2006). TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. 46: 768–779 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
