

Viral interactions with B-cells contribute to increased regulatory T-cells during chronic HCV infection
- PMID: 21449722
- PMCID: PMC3070003
- DOI: 10.1089/vim.2010.0077
Viral interactions with B-cells contribute to increased regulatory T-cells during chronic HCV infection
Abstract
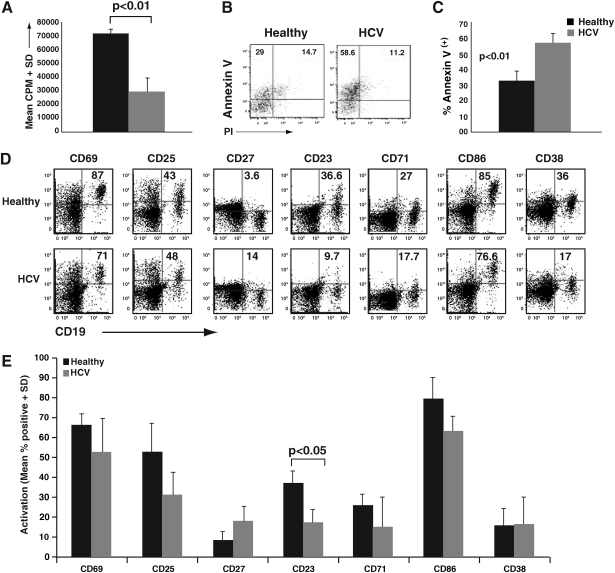
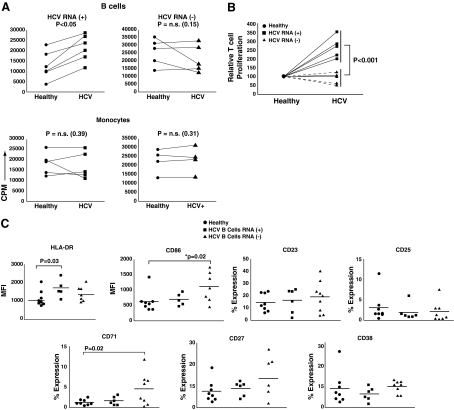
Hepatitis C virus (HCV) has a propensity to establish chronic infection that is characterized by attenuated virus-specific T-cell responses. Mechanisms leading to T-cell attenuation are poorly understood and likely involve dysfunctional interactions between antigen-presenting cells (APC) and effector/regulatory T-cells. Reports on dendritic cells (DC) have described only minor dysfunction during HCV infection. However, there is a paucity of reports regarding B-cell function, despite clear associations with B-cell-related secondary sequelae. In this study we evaluated the state of B-cells during chronic HCV infection, and observed a diminished ability to respond to mitogenic stimuli, correlating with increased apoptosis. This was in contrast to their ex vivo phenotype, which indicated ongoing chronic activation in vivo. There was a high association of HCV-positive strand RNA with B-cells in a subset of HCV patients. Interestingly, ex-vivo-derived HCV RNA-positive B-cells induced significantly greater proliferation in allogeneic T-cells than in HCV-negative B-cells, correlating with an increased generation of CD4(+)CD25(+)FOXP3(+) regulatory T-cells (Tregs). In-vitro exposure of healthy peripheral blood mononuclear cells (PBMC) to HCV resulted in robust activation of resting B-cells. These HCV-exposed B-cells also showed an enhanced ability to generate Tregs. Our results provide strong evidence for a novel and paradoxical link between HCV-induced enhanced APC function and the generation of Tregs.
Figures


Similar articles
-
Identification and in vitro expansion of functional antigen-specific CD25+ FoxP3+ regulatory T cells in hepatitis C virus infection.J Virol. 2008 May;82(10):5043-53. doi: 10.1128/JVI.01548-07. Epub 2008 Mar 12. J Virol. 2008. PMID: 18337568 Free PMC article.
-
Defining target antigens for CD25+ FOXP3 + IFN-gamma- regulatory T cells in chronic hepatitis C virus infection.Immunol Cell Biol. 2007 Apr-May;85(3):197-204. doi: 10.1038/sj.icb.7100020. Epub 2007 Jan 2. Immunol Cell Biol. 2007. PMID: 17199111
-
F protein increases CD4+CD25+ T cell population in patients with chronic hepatitis C.Pathog Dis. 2015 Jun;73(4):ftv022. doi: 10.1093/femspd/ftv022. Epub 2015 Apr 9. Pathog Dis. 2015. PMID: 25862675
-
Dendritic cells, regulatory T cells and the pathogenesis of chronic hepatitis C.Virulence. 2012 Nov 15;3(7):610-20. doi: 10.4161/viru.21823. Epub 2012 Oct 17. Virulence. 2012. PMID: 23076334 Free PMC article. Review.
-
Divergent contributions of regulatory T cells to the pathogenesis of chronic hepatitis C.Hum Vaccin Immunother. 2013 Jul;9(7):1569-76. doi: 10.4161/hv.24726. Epub 2013 May 31. Hum Vaccin Immunother. 2013. PMID: 23732899 Free PMC article. Review.
Cited by
-
CD19+CD24hiCD38hi regulatory B cells: a potential immune predictive marker of severity and therapeutic responsiveness of hepatitis C.Am J Transl Res. 2020 Mar 15;12(3):889-900. eCollection 2020. Am J Transl Res. 2020. PMID: 32269721 Free PMC article.
-
Modulation of immune function occurs within hours of therapy initiation for multiple sclerosis.Clin Immunol. 2013 May;147(2):105-19. doi: 10.1016/j.clim.2013.02.015. Epub 2013 Mar 6. Clin Immunol. 2013. PMID: 23578552 Free PMC article. Clinical Trial.
-
Effects of Friend Virus Infection and Regulatory T Cells on the Antigen Presentation Function of B Cells.mBio. 2019 Jan 22;10(1):e02578-18. doi: 10.1128/mBio.02578-18. mBio. 2019. PMID: 30670616 Free PMC article.
-
Detection of hepatitis C virus (HCV) negative strand RNA and NS3 protein in peripheral blood mononuclear cells (PBMC): CD3+, CD14+ and CD19+.Virol J. 2013 Nov 26;10:346. doi: 10.1186/1743-422X-10-346. Virol J. 2013. PMID: 24279719 Free PMC article.
References
-
- Altomonte M. Montagner R. Pucillo C. Maio M. Triggering of target of an antiproliferative antibody-1 (TAPA-1/CD81) up-regulates the release of tumour necrosis factor-alpha by the EBV-B lymphoblastoid cell line JY. Scand J Immunol. 1996;43:367–373. - PubMed
-
- Bain C. Fatmi A. Zoulim F, et al. Impaired allostimulatory function of dendritic cells in chronic hepatitis C infection. Gastroenterology. 2001;120:512–524. - PubMed
-
- Barnes E. Lauer G. Walker B. Klenerman P. T cell failure in hepatitis C virus infection. Viral Immunol. 2002;15:285–293. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
