

Plio-Pleistocene sea level and temperature fluctuations in the northwestern Pacific promoted speciation in the globally-distributed flathead mullet Mugil cephalus
- PMID: 21450111
- PMCID: PMC3079632
- DOI: 10.1186/1471-2148-11-83
Plio-Pleistocene sea level and temperature fluctuations in the northwestern Pacific promoted speciation in the globally-distributed flathead mullet Mugil cephalus
Abstract
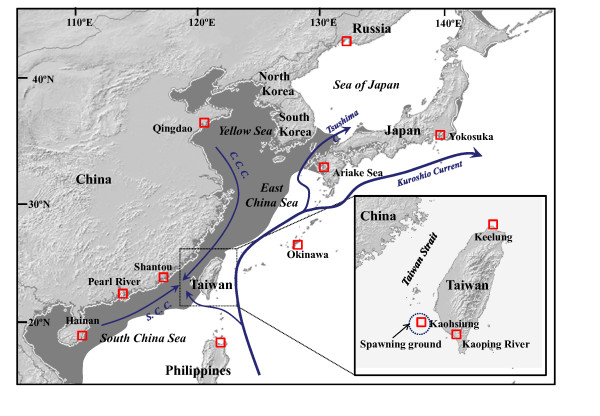
Background: The study of speciation in the marine realm is challenging because of the apparent absence of physical barriers to dispersal, which are one of the main drivers of genetic diversity. Although phylogeographic studies using mitochondrial DNA (mtDNA) information often reveal significant genetic heterogeneity within marine species, the evolutionary significance of such diversity is difficult to interpret with these markers. In the northwestern (NW) Pacific, several studies have emphasised the potential importance of sea-level regression during the most recent glaciations as a driver of genetic diversity in marine species. These studies have failed, however, to determine whether the period of isolation was long enough for divergence to attain speciation. Among these marine species, the cosmopolitan estuarine-dependent fish Mugil cephalus represents an interesting case study. Several divergent allopatric mtDNA lineages have been described in this species worldwide, and three occur in sympatry in the NW Pacific.
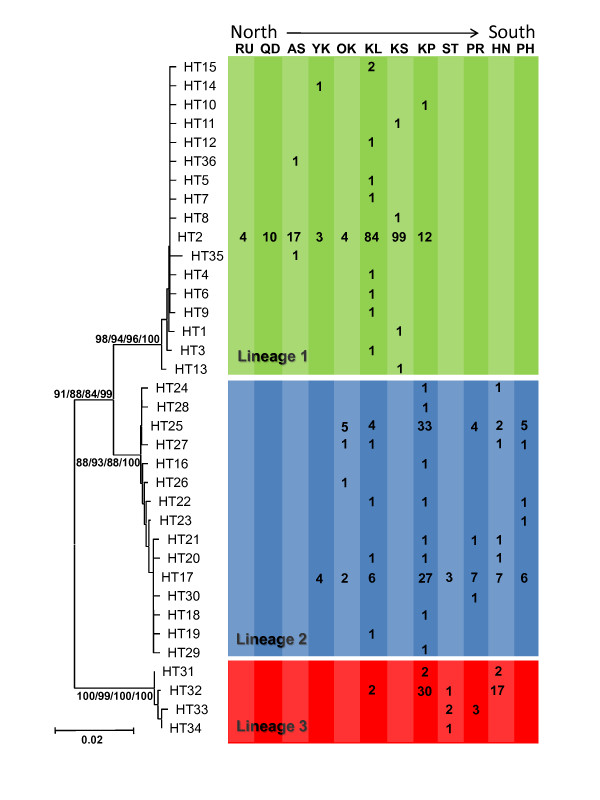
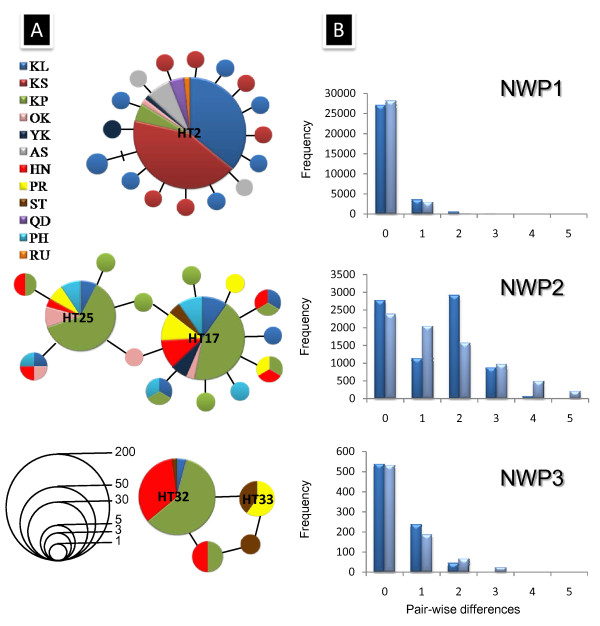
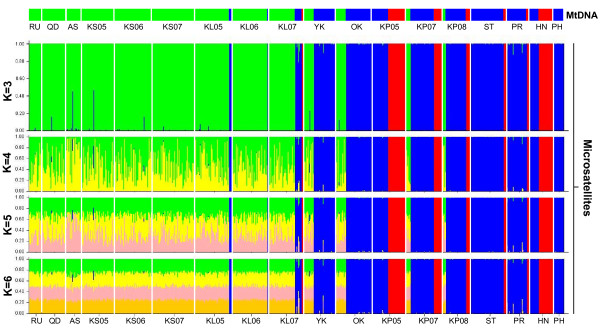
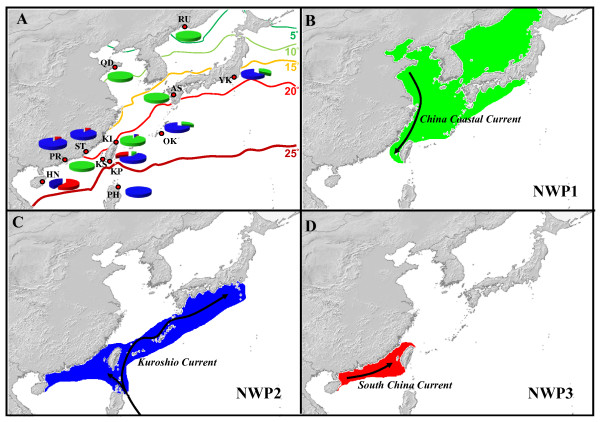
Results: Ten nuclear microsatellites were surveyed to estimate the level of genetic isolation of these lineages and determine the role of sea-level fluctuation in the evolution of NW Pacific M. cephalus. Three cryptic species of M. cephalus were identified within this region (NWP1, 2 and 3) using an assignment test on the microsatellite data. Each species corresponds with one of the three mtDNA lineages in the COI phylogenetic tree. NWP3 is the most divergent species, with a distribution range that suggests tropical affinities, while NWP1, with a northward distribution from Taiwan to Russia, is a temperate species. NWP2 is distributed along the warm Kuroshio Current. The divergence of NWP1 from NWP2 dates back to the Pleistocene epoch and probably corresponds to the separation of the Japan and China Seas when sea levels dropped. Despite their subsequent range expansion since this period of glaciation, no gene flow was observed among these three lineages, indicating that speciation has been achieved.
Conclusions: This study successfully identified three cryptic species in M. cephalus inhabiting the NW Pacific, using a combination of microsatellites and mitochondrial genetic markers. The current genetic architecture of the M. cephalus species complex in the NW Pacific is the result of a complex interaction of contemporary processes and historical events. Sea level and temperature fluctuations during Plio-Pleistocene epochs probably played a major role in creating the marine species diversity of the NW Pacific that is found today.
Figures
References
-
- Haig JA, Connolly RM, Hughes JM. Little shrimp left on the shelf: the roles that sea-level change, ocean currents and continental shelf width play in the genetic connectivity of a seagrass-associated species. J Biogeog. 2010;37:1570–1583.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
