

Both visual and idiothetic cues contribute to head direction cell stability during navigation along complex routes
- PMID: 21451060
- PMCID: PMC3118751
- DOI: 10.1152/jn.01041.2010
Both visual and idiothetic cues contribute to head direction cell stability during navigation along complex routes
Abstract
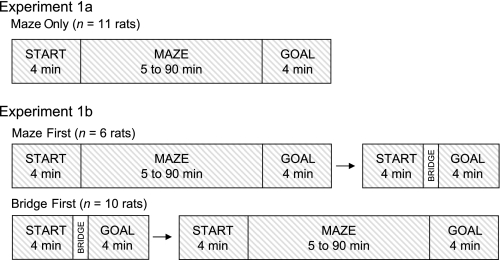
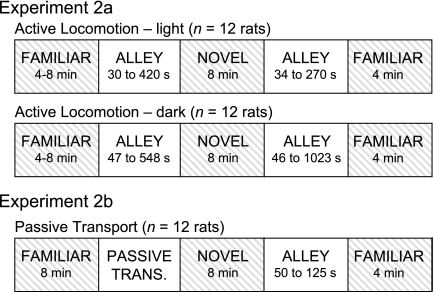
Successful navigation requires a constantly updated neural representation of directional heading, which is conveyed by head direction (HD) cells. The HD signal is predominantly controlled by visual landmarks, but when familiar landmarks are unavailable, self-motion cues are able to control the HD signal via path integration. Previous studies of the relationship between HD cell activity and path integration have been limited to two or more arenas located in the same room, a drawback for interpretation because the same visual cues may have been perceptible across arenas. To address this issue, we tested the relationship between HD cell activity and path integration by recording HD cells while rats navigated within a 14-unit T-maze and in a multiroom maze that consisted of unique arenas that were located in different rooms but connected by a passageway. In the 14-unit T-maze, the HD signal remained relatively stable between the start and goal boxes, with the preferred firing directions usually shifting <45° during maze traversal. In the multiroom maze in light, the preferred firing directions also remained relatively constant between rooms, but with greater variability than in the 14-unit maze. In darkness, HD cell preferred firing directions showed marginally more variability between rooms than in the lighted condition. Overall, the results indicate that self-motion cues are capable of maintaining the HD cell signal in the absence of familiar visual cues, although there are limits to its accuracy. In addition, visual information, even when unfamiliar, can increase the precision of directional perception.
Figures








References
-
- Batschelet E. Circular Statistics in Biology. New York: Academic, 1981
-
- Benhamou S, Sauvé JP, Bovet P. Spatial memory in large-scale environments: efficiency and limitations of the egocentric coding process. J Theor Biol 145: 1–12, 1990
-
- Bingman VP, Cheng K. Mechanisms of animal global navigation: comparative perspectives and enduring challenges. Ethol Ecol Evol 17: 295–318, 2005
-
- Blair HT, Sharp PE. Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat. Behav Neurosci 110: 643–660, 1996 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources