

Similar mitochondrial activation kinetics in wild-type and creatine kinase-deficient fast-twitch muscle indicate significant Pi control of respiration
- PMID: 21451138
- PMCID: PMC3119146
- DOI: 10.1152/ajpregu.00204.2010
Similar mitochondrial activation kinetics in wild-type and creatine kinase-deficient fast-twitch muscle indicate significant Pi control of respiration
Abstract
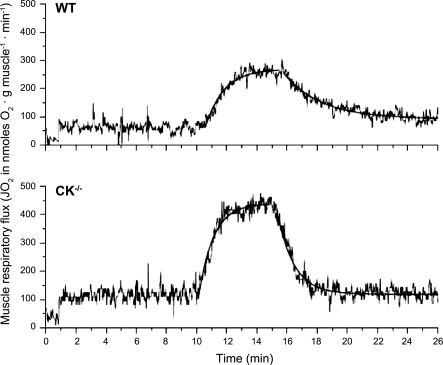
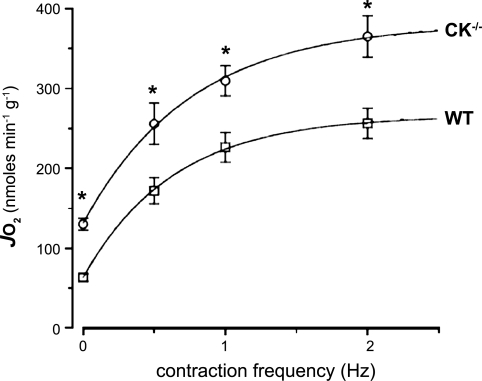
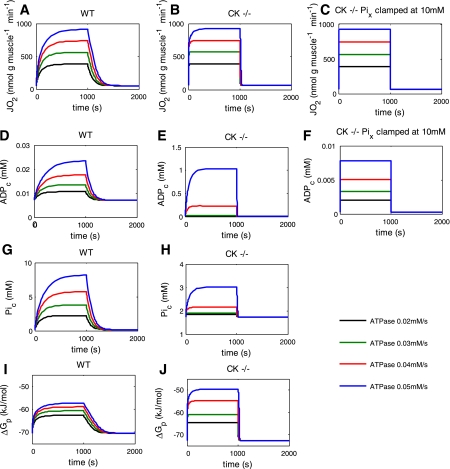
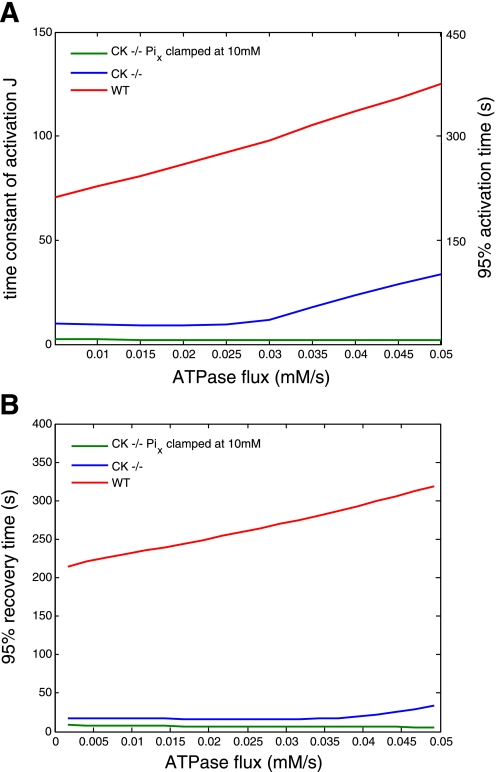
Past simulations of oxidative ATP metabolism in skeletal muscle have predicted that elimination of the creatine kinase (CK) reaction should result in dramatically faster oxygen consumption dynamics during transitions in ATP turnover rate. This hypothesis was investigated. Oxygen consumption of fast-twitch (FT) muscle isolated from wild-type (WT) and transgenic mice deficient in the myoplasmic (M) and mitochondrial (Mi) CK isoforms (MiM CK(-/-)) were measured at 20°C at rest and during electrical stimulation. MiM CK(-/-) muscle oxygen consumption activation kinetics during a step change in contraction rate were 30% faster than WT (time constant 53 ± 3 vs. 69 ± 4 s, respectively; mean ± SE, n = 8 and 6, respectively). MiM CK(-/-) muscle oxygen consumption deactivation kinetics were 380% faster than WT (time constant 74 ± 4 s vs. 264 ± 4 s, respectively). Next, the experiments were simulated using a computational model of the oxidative ATP metabolic network in FT muscle featuring ADP and Pi feedback control of mitochondrial respiration (J. A. L. Jeneson, J. P. Schmitz, N. A. van den Broek, N. A. van Riel, P. A. Hilbers, K. Nicolay, J. J. Prompers. Am J Physiol Endocrinol Metab 297: E774-E784, 2009) that was reparameterized for 20°C. Elimination of Pi control via clamping of the mitochondrial Pi concentration at 10 mM reproduced past simulation results of dramatically faster kinetics in CK(-/-) muscle, while inclusion of Pi control qualitatively explained the experimental observations. On this basis, it was concluded that previous studies of the CK-deficient FT muscle phenotype underestimated the contribution of Pi to mitochondrial respiratory control.
Figures
References
-
- Bose S, French S, Evans FJ, Joubert F, Balaban RS. Metabolic network control of oxidative phosphorylation: multiple roles of inorganic phosphate. J Biol Chem 278: 39155–39165, 2003 - PubMed
-
- Brand MD. The efficiency and plasticity of mitochondrial energy transduction. Biochem Soc Trans 33: 897–904, 2005 - PubMed
-
- Chance B, Im J, Nioka S, Kushmerick MJ. Skeletal muscle energetics with PNMR: personal views and historic perspectives. NMR Biomed 19: 904–926, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous