

MRB3010 is a core component of the MRB1 complex that facilitates an early step of the kinetoplastid RNA editing process
- PMID: 21451155
- PMCID: PMC3078736
- DOI: 10.1261/rna.2446311
MRB3010 is a core component of the MRB1 complex that facilitates an early step of the kinetoplastid RNA editing process
Abstract
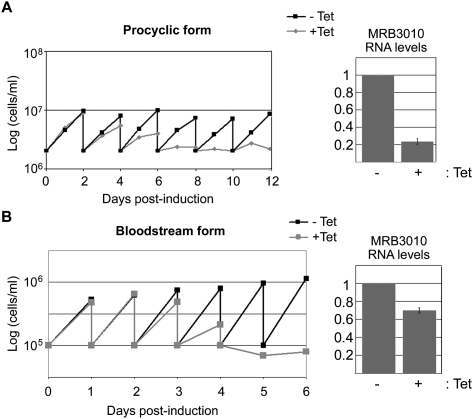
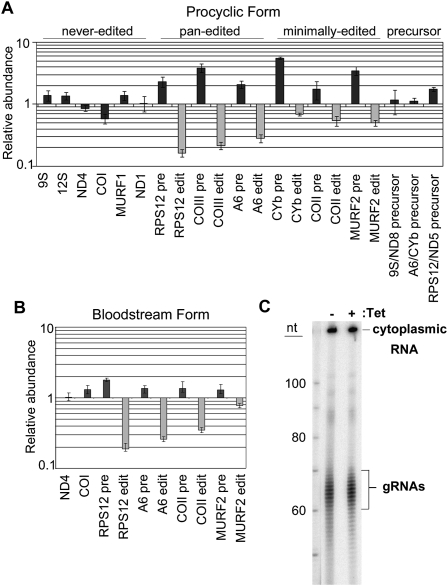
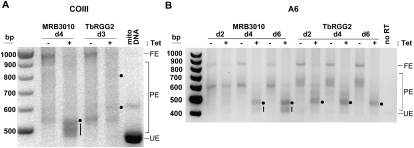
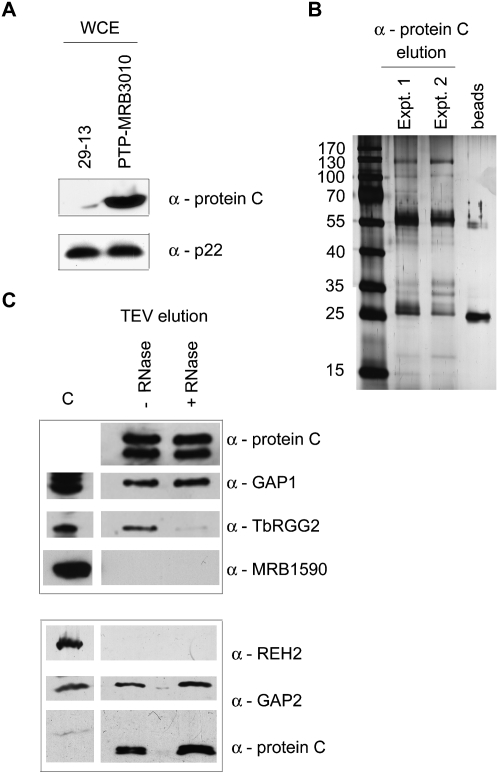
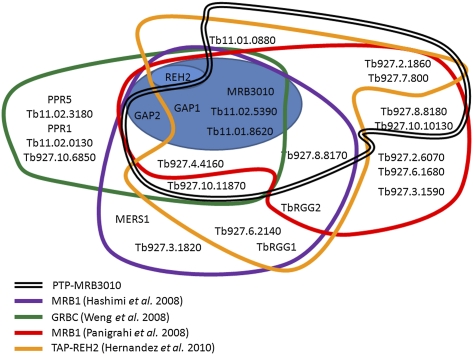
Gene expression in the mitochondria of the kinetoplastid parasite Trypanosoma brucei is regulated primarily post-transcriptionally at the stages of RNA processing, editing, and turnover. The mitochondrial RNA-binding complex 1 (MRB1) is a recently identified multiprotein complex containing components with distinct functions during different aspects of RNA metabolism, such as guide RNA (gRNA) and mRNA turnover, precursor transcript processing, and RNA editing. In this study we examined the function of the MRB1 protein, Tb927.5.3010, which we term MRB3010. We show that MRB3010 is essential for growth of both procyclic form and bloodstream form life-cycle stages of T. brucei. Down-regulation of MRB3010 by RNAi leads to a dramatic inhibition of RNA editing, yet its depletion does not impact total gRNA levels. Rather, it appears to affect the editing process at an early stage, as indicated by the accumulation of pre-edited and small partially edited RNAs. MRB3010 is present in large (>20S) complexes and exhibits both RNA-dependent and RNA-independent interactions with other MRB1 complex proteins. Comparison of proteins isolated with MRB3010 tagged at its endogenous locus to those reported from other MRB1 complex purifications strongly suggests the presence of an MRB1 "core" complex containing five to six proteins, including MRB3010. Together, these data further our understanding of the function and composition of the imprecisely defined MRB1 complex.
Figures
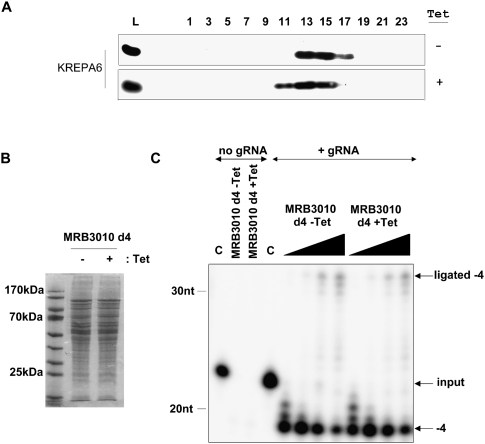
 -32P]ATP-radiolabeled 5′ mRNA fragment and 3′ mRNA fragment, but no gRNA (no gRNA); or radiolabeled 5′ mRNA fragment, 3′ mRNA fragment, and gRNA (+ gRNA). In the no gRNA experiments either a control with no protein (C) or 2.5 μL of the indicated extract was added. Nonspecific ribonucleases in the extracts degraded the radiolabeled 5′ mRNA fragment when gRNA was not present. In the experiments where gRNA was added either no protein (C) or 1, 2.5, 5, and 7.5 μL of the indicated extracts were added. The migration position of the input radiolabeled 5′ mRNA fragment is indicated. The −4 nonligated deletion product is labeled −4 and the ligated deletion product is labeled ligated −4.
-32P]ATP-radiolabeled 5′ mRNA fragment and 3′ mRNA fragment, but no gRNA (no gRNA); or radiolabeled 5′ mRNA fragment, 3′ mRNA fragment, and gRNA (+ gRNA). In the no gRNA experiments either a control with no protein (C) or 2.5 μL of the indicated extract was added. Nonspecific ribonucleases in the extracts degraded the radiolabeled 5′ mRNA fragment when gRNA was not present. In the experiments where gRNA was added either no protein (C) or 1, 2.5, 5, and 7.5 μL of the indicated extracts were added. The migration position of the input radiolabeled 5′ mRNA fragment is indicated. The −4 nonligated deletion product is labeled −4 and the ligated deletion product is labeled ligated −4.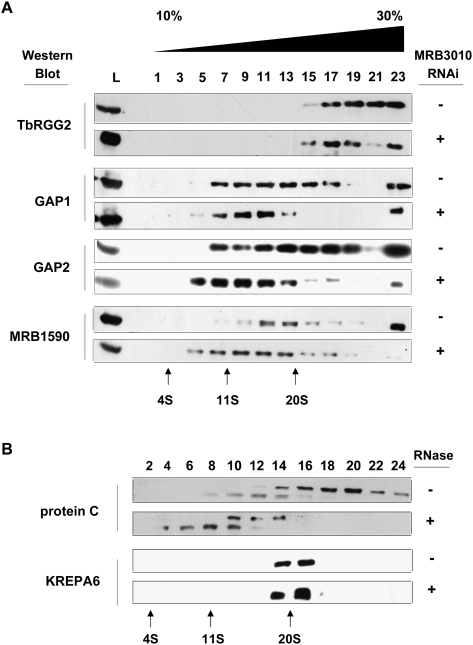
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases