

Fezf1 and Fezf2 are required for olfactory development and sensory neuron identity
- PMID: 21452247
- PMCID: PMC3268373
- DOI: 10.1002/cne.22596
Fezf1 and Fezf2 are required for olfactory development and sensory neuron identity
Abstract
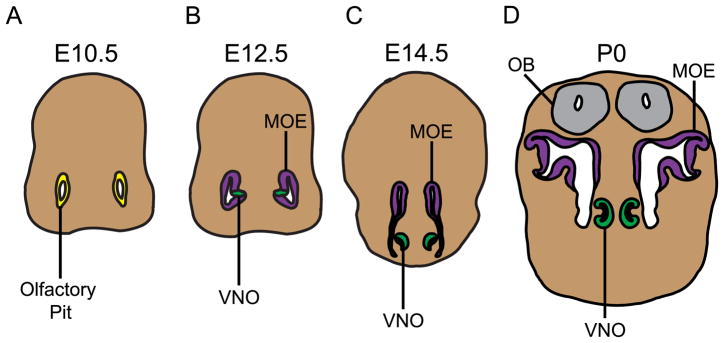
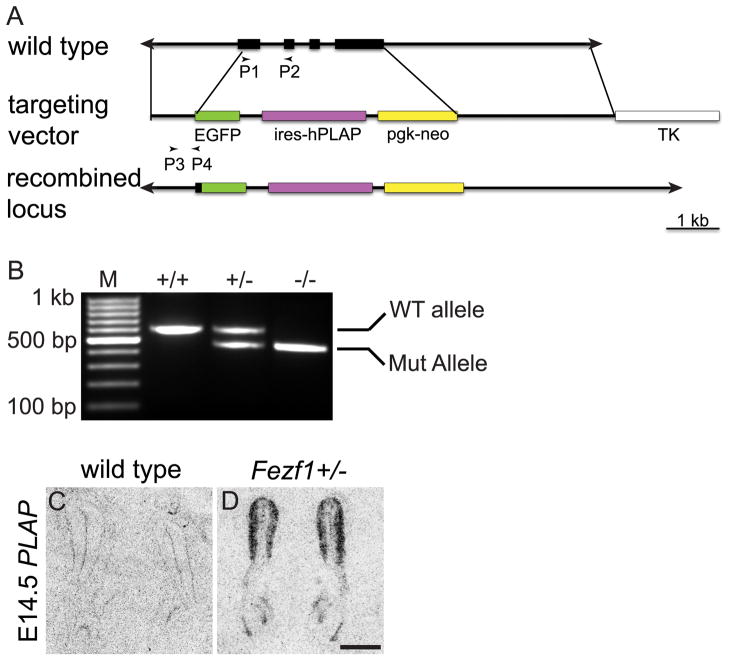
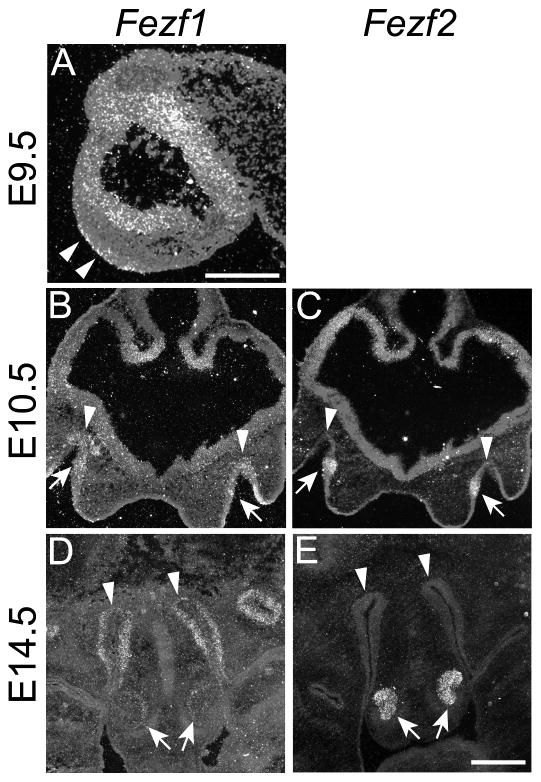
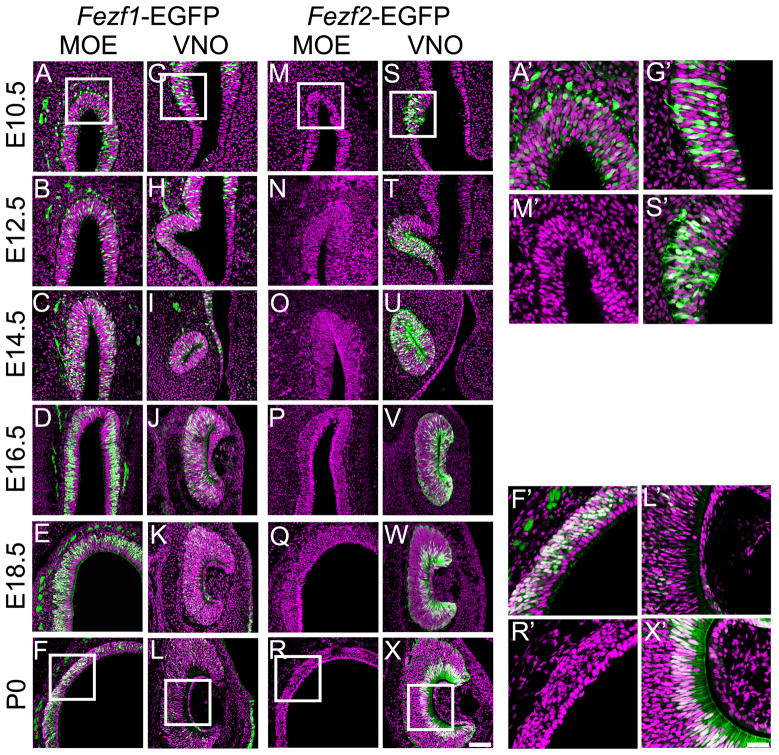
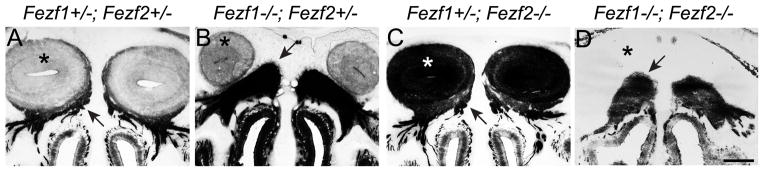
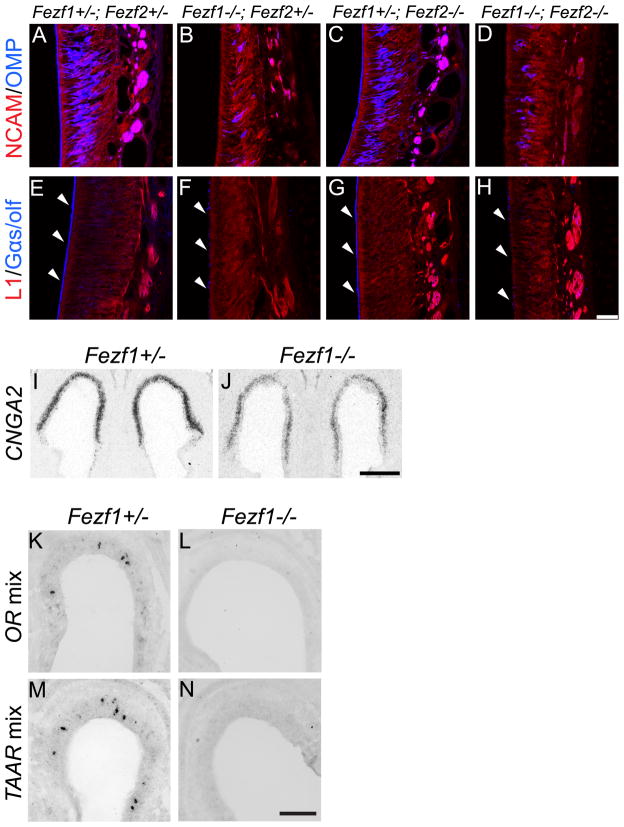
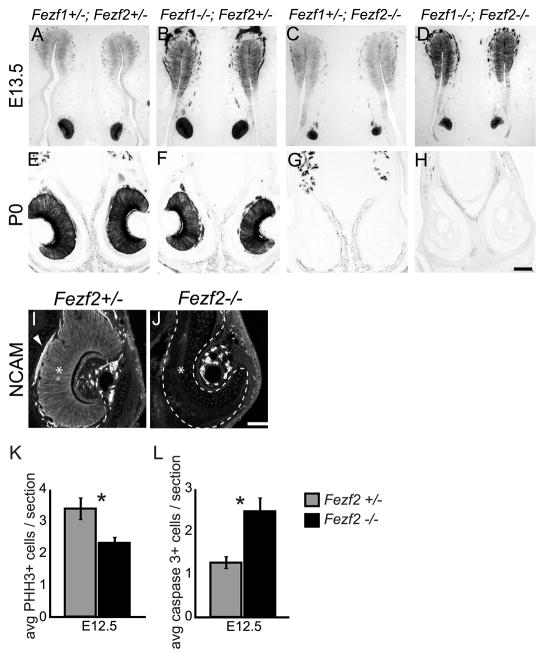
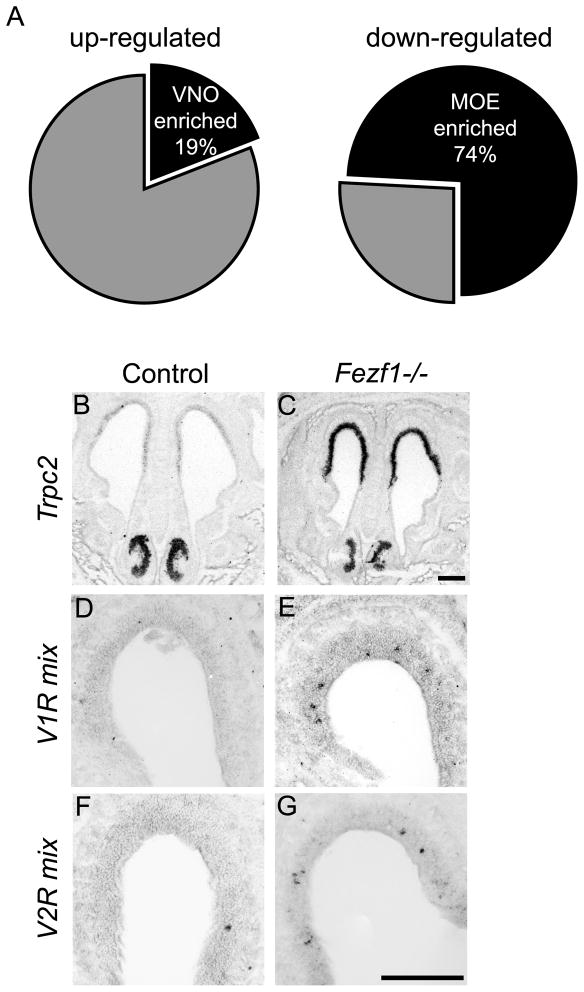
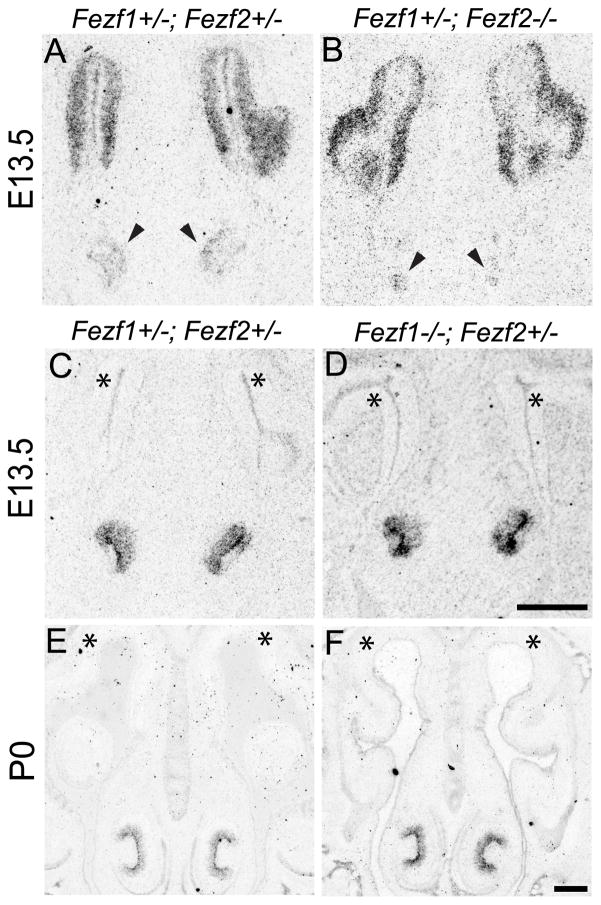
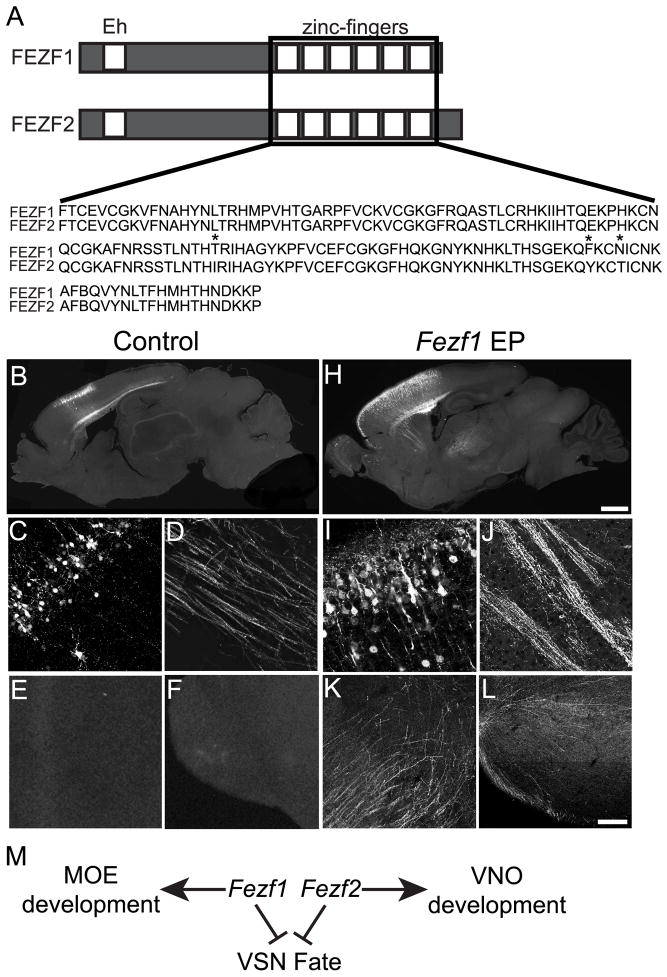
The murine olfactory system consists of main and accessory systems that perform distinct and overlapping functions. The main olfactory epithelium (MOE) is primarily involved in the detection of volatile odorants, while neurons in the vomeronasal organ (VNO), part of the accessory olfactory system, are important for pheromone detection. During development, the MOE and VNO both originate from the olfactory pit; however, the mechanisms regulating development of these anatomically distinct organs from a common olfactory primordium are unknown. Here we report that two closely related zinc-finger transcription factors, FEZF1 and FEZF2, regulate the identity of MOE sensory neurons and are essential for the survival of VNO neurons respectively. Fezf1 is predominantly expressed in the MOE while Fezf2 expression is restricted to the VNO. In Fezf1-deficient mice, olfactory neurons fail to mature and also express markers of functional VNO neurons. In Fezf2-deficient mice, VNO neurons degenerate prior to birth. These results identify Fezf1 and Fezf2 as important regulators of olfactory system development and sensory neuron identity.
Copyright © 2011 Wiley-Liss, Inc.
Figures
References
-
- Baker H, Grillo M, Margolis FL. Biochemical and immunocytochemical characterization of olfactory marker protein in the rodent central nervous system. J Comp Neurol. 1989;285(2):246–261. - PubMed
-
- Balmer CW, LaMantia AS. Noses and neurons: induction, morphogenesis, and neuronal differentiation in the peripheral olfactory pathway. Dev Dyn. 2005;234(3):464–481. - PubMed
-
- Beites CL, Kawauchi S, Crocker CE, Calof AL. Identification and molecular regulation of neural stem cells in the olfactory epithelium. Exp Cell Res. 2005;306(2):309–316. - PubMed
-
- Cau E, Casarosa S, Guillemot F. Mash1 and Ngn1 control distinct steps of determination and differentiation in the olfactory sensory neuron lineage. Development. 2002;129(8):1871–1880. - PubMed
-
- Cau E, Gradwohl G, Casarosa S, Kageyama R, Guillemot F. Hes genes regulate sequential stages of neurogenesis in the olfactory epithelium. Development. 2000;127(11):2323–2332. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
