

The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype
- PMID: 21453429
- PMCID: PMC6640367
- DOI: 10.1111/j.1364-3703.2010.00674.x
The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype
Abstract
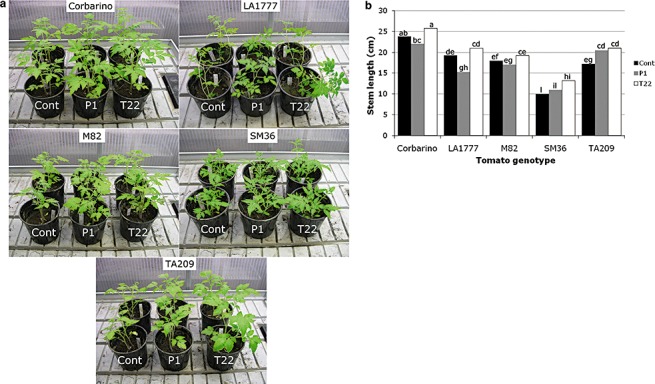
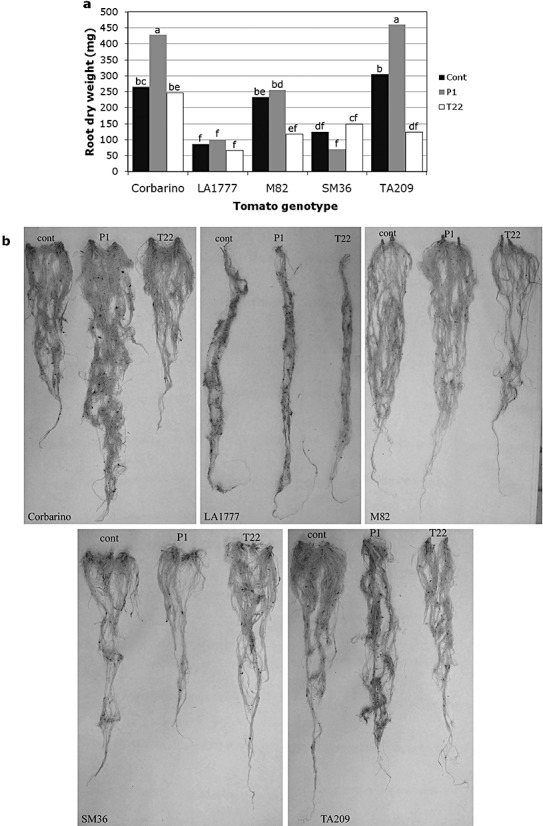
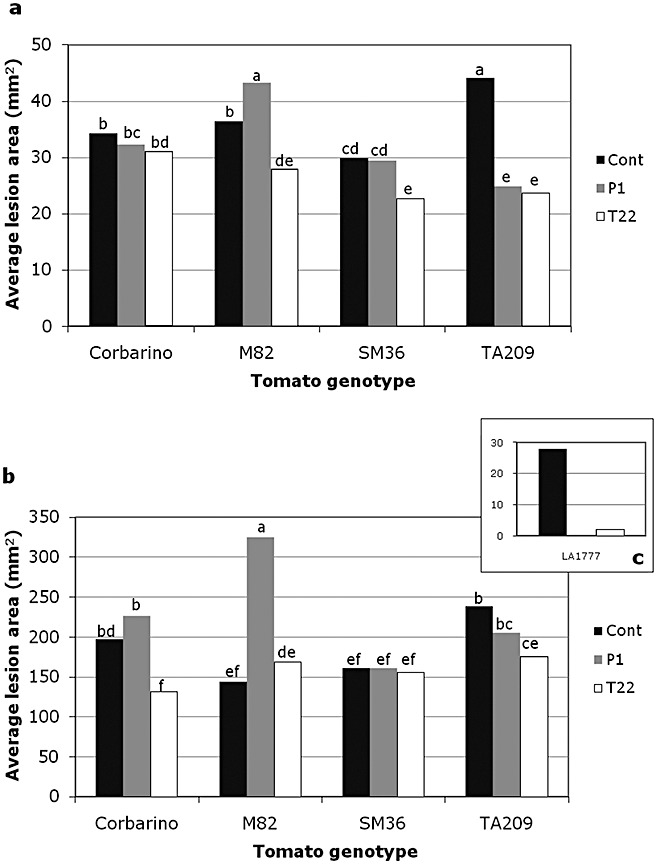
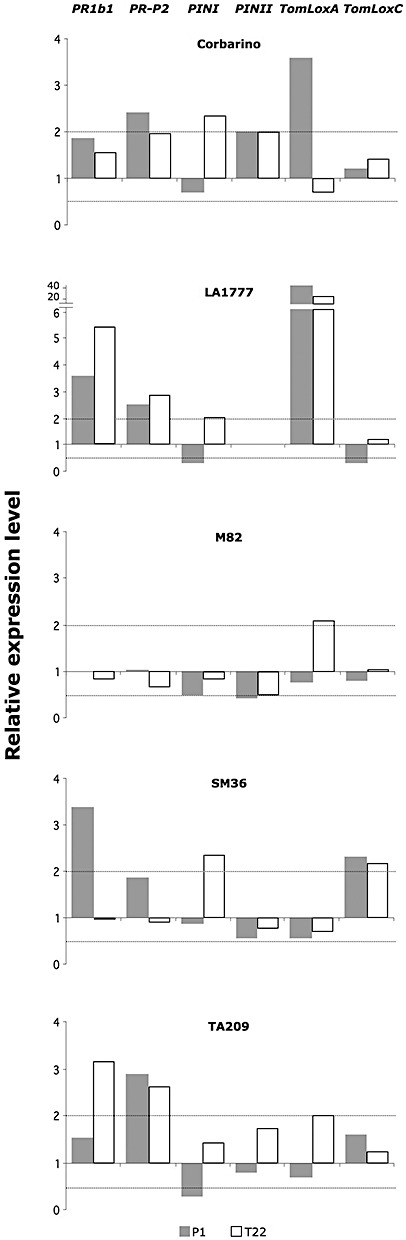
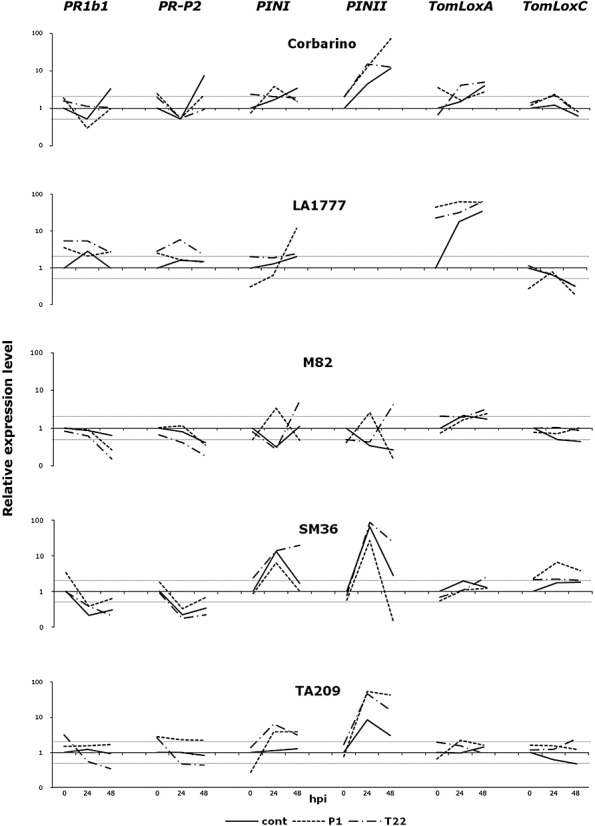
Rhizosphere-competent fungi of the genus Trichoderma are widely used as biofertilizers and biopesticides in commercial formulates because of the multiple beneficial effects on plant growth and disease resistance. In this work, we demonstrate that genetic variability among wild and cultivated tomato lines affects the outcome of the interaction with two 'elite' biocontrol strains of T. atroviride and T. harzianum. The beneficial response, which included enhanced growth and systemic resistance against Botrytis cinerea, was clearly evident for some, but not all, the tested lines. At least in one case (line M82), treatment with the biocontrol agents had no effect or was even detrimental. Expression studies on defence-related genes suggested that the fungus is able to trigger, in the responsive lines, a long-lasting up-regulation of the salicylic acid pathway in the absence of a pathogen, possibly activating a priming mechanism in the plant. Consequently, infection with B. cinerea on plants pretreated with Trichoderma is followed by enhanced activation of jasmonate-responsive genes, eventually boosting systemic resistance to the pathogen in a plant genotype-dependent manner. Our data indicate that, at least in tomato, the Trichoderma induced systemic resistance mechanism is much more complex than considered so far, and the ability of the plant to benefit from this symbiotic-like interaction can be genetically improved.
MOLECULAR PLANT PATHOLOGY © 2010 BSPP AND BLACKWELL PUBLISHING LTD. NO CLAIM TO ORIGINAL US GOVERNMENT WORKS.
Figures
References
-
- Ahn, I.P. , Lee, S.W. and Suh, S.C. (2007) Rhizobacteria‐induced priming in Arabidopsis is dependent on ethylene, jasmonic acid, and NPR1. Mol. Plant–Microbe Interact. 20, 759–768. - PubMed
-
- Alfano, G. , Ivey, M.L. , Cakir, C. , Bos, J.I. , Miller, S.A. , Madden, L.V. , Kamoun, S. and Hoitink, H.A. (2007) Systemic modulation of gene expression in tomato by Trichoderma hamatum 382. Phytopathology, 97, 429–437. - PubMed
-
- Benitez, T. , Rincon, A.M. , Limon, M.C. and Codon, A.C. (2004) Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 7, 249–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
