

N-acetylaspartylglutamate synthetase II synthesizes N-acetylaspartylglutamylglutamate
- PMID: 21454531
- PMCID: PMC3089511
- DOI: 10.1074/jbc.M111.230136
N-acetylaspartylglutamate synthetase II synthesizes N-acetylaspartylglutamylglutamate
Abstract
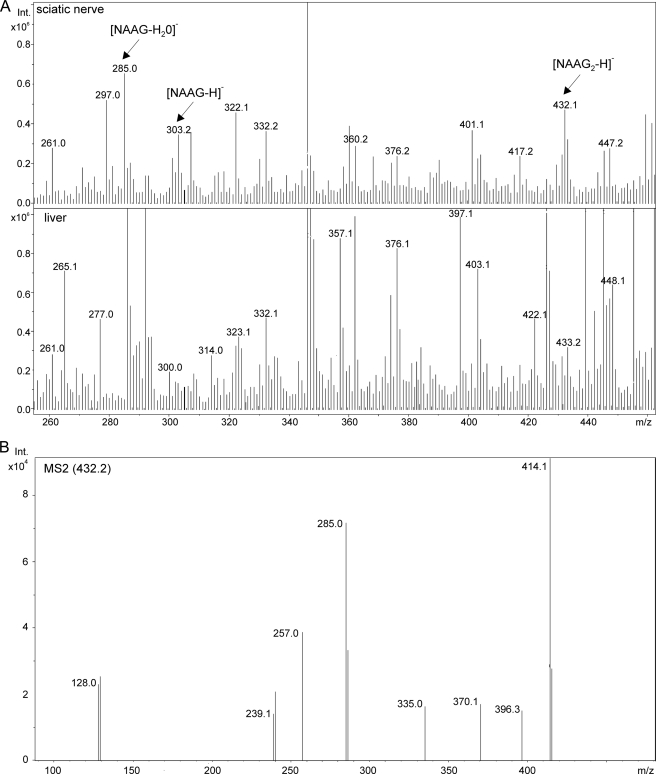
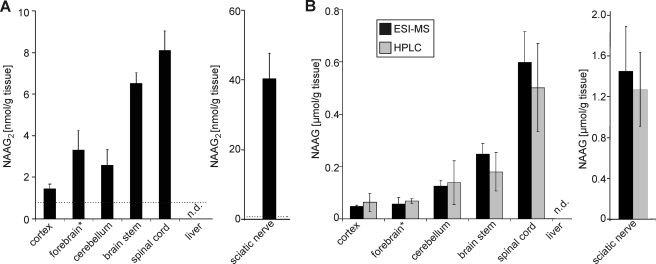
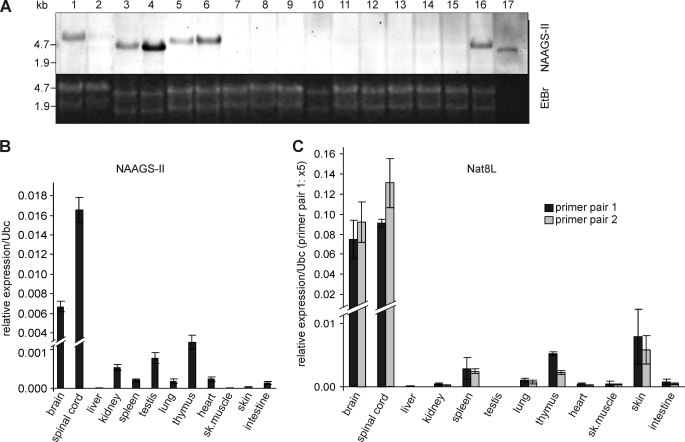
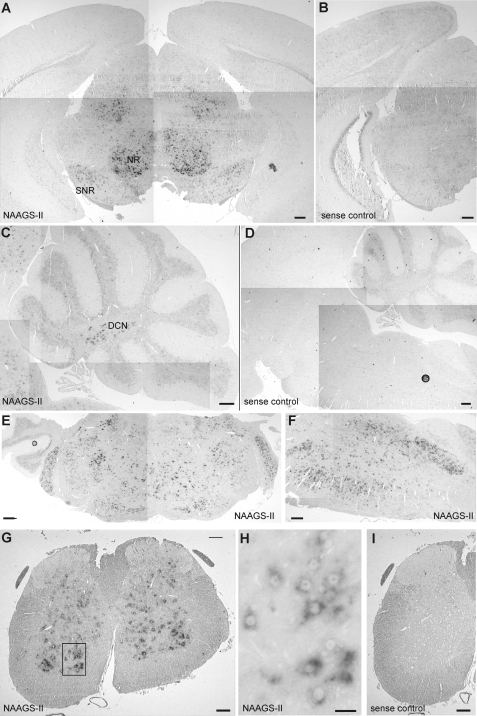
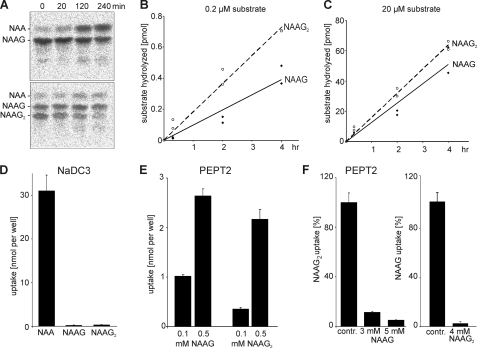
N-Acetylaspartylglutamate (NAAG) is found at high concentrations in the vertebrate nervous system. NAAG is an agonist at group II metabotropic glutamate receptors. In addition to its role as a neuropeptide, a number of functions have been proposed for NAAG, including a role as a non-excitotoxic transport form of glutamate and a molecular water pump. We recently identified a NAAG synthetase (now renamed NAAG synthetase I, NAAGS-I), encoded by the ribosomal modification protein rimK-like family member B (Rimklb) gene, as a member of the ATP-grasp protein family. We show here that a structurally related protein, encoded by the ribosomal modification protein rimK-like family member A (Rimkla) gene, is another NAAG synthetase (NAAGS-II), which in addition, synthesizes the N-acetylated tripeptide N-acetylaspartylglutamylglutamate (NAAG(2)). In contrast, NAAG(2) synthetase activity was undetectable in cells expressing NAAGS-I. Furthermore, we demonstrate by mass spectrometry the presence of NAAG(2) in murine brain tissue and sciatic nerves. The highest concentrations of both, NAAG(2) and NAAG, were found in sciatic nerves, spinal cord, and the brain stem, in accordance with the expression level of NAAGS-II. To our knowledge the presence of NAAG(2) in the vertebrate nervous system has not been described before. The physiological role of NAAG(2), e.g. whether it acts as a neurotransmitter, remains to be determined.
Figures

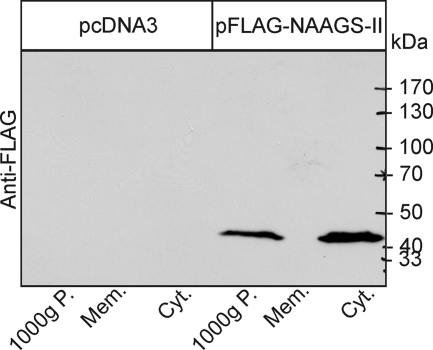
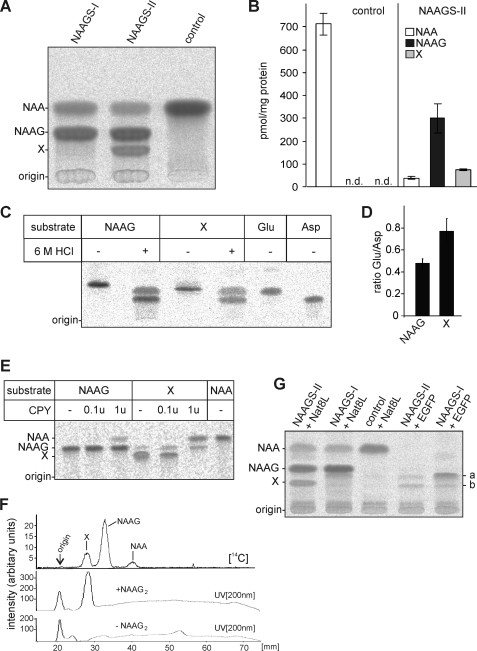
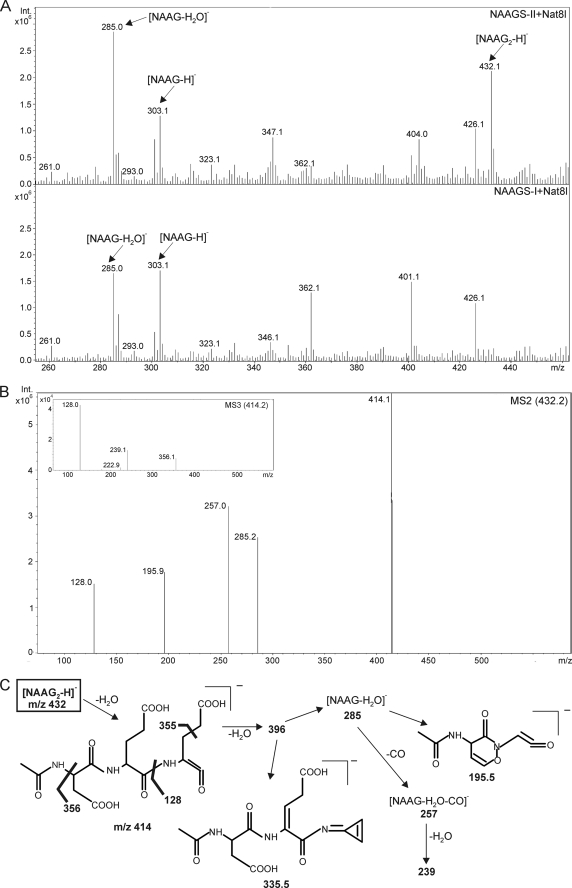
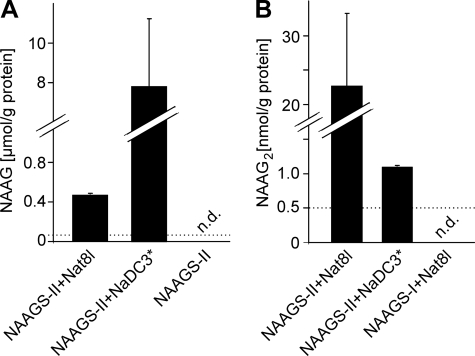
References
-
- Curatolo A., D'Arcangelo P., Lino A., Brancati A. (1965) J. Neurochem. 12, 339–342 - PubMed
-
- Coyle J. T. (1997) Neurobiol. Dis. 4, 231–238 - PubMed
-
- Neale J. H., Bzdega T., Wroblewska B. (2000) J. Neurochem. 75, 443–452 - PubMed
-
- Wroblewska B., Wroblewski J. T., Pshenichkin S., Surin A., Sullivan S. E., Neale J. H. (1997) J. Neurochem. 69, 174–181 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
