

Single substitution within the RKTR motif impairs kinase activity but promotes dimerization of RAF kinase
- PMID: 21454547
- PMCID: PMC3091254
- DOI: 10.1074/jbc.M110.194167
Single substitution within the RKTR motif impairs kinase activity but promotes dimerization of RAF kinase
Abstract
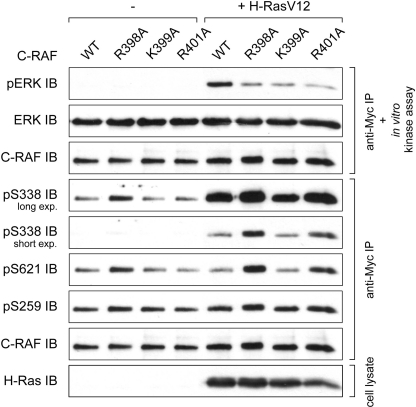
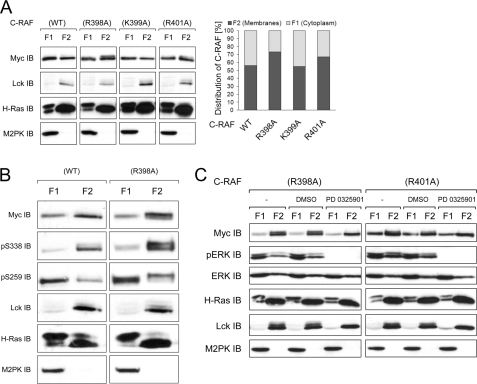
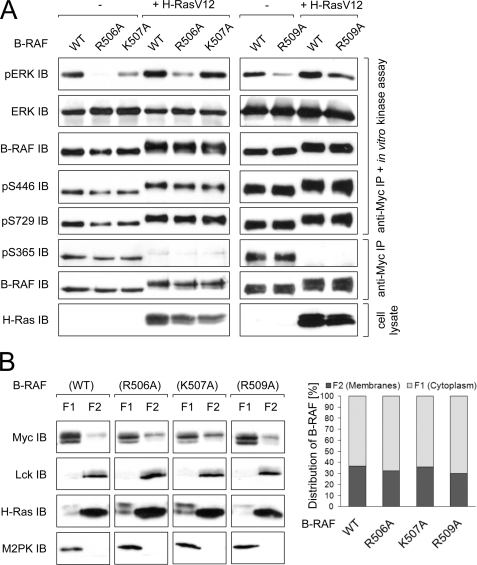
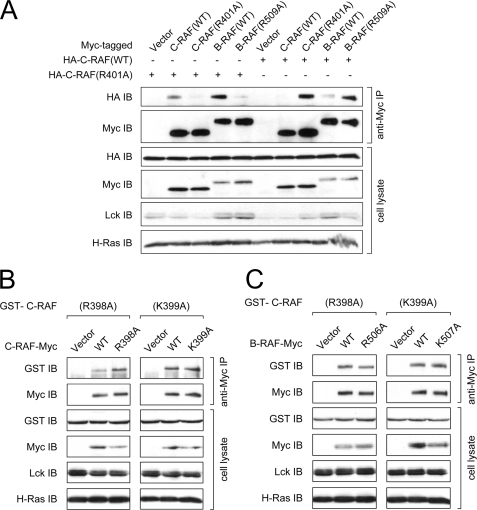
The serine/threonine kinase RAF is a central component of the MAPK cascade. Regulation of RAF activity is highly complex and involves recruitment to membranes and association with Ras and scaffold proteins as well as multiple phosphorylation and dephosphorylation events. Previously, we identified by molecular modeling an interaction between the N-region and the RKTR motif of the kinase domain in RAF and assigned a new function to this tetrapeptide segment. Here we found that a single substitution of each basic residue within the RKTR motif inhibited catalytic activity of all three RAF isoforms. However, the inhibition and phosphorylation pattern of C-RAF and A-RAF differed from B-RAF. Furthermore, substitution of the first arginine led to hyperphosphorylation and accumulation of A-RAF and C-RAF in plasma membrane fraction, indicating that this residue interferes with the recycling process of A-RAF and C-RAF but not B-RAF. In contrast, all RAF isoforms behave similarly with respect to the RKTR motif-dependent dimerization. The exchange of the second arginine led to exceedingly increased dimerization as long as one of the protomers was not mutated, suggesting that substitution of this residue with alanine may result in similar a structural rearrangement of the RAF kinase domain, as has been found for the C-RAF kinase domain co-crystallized with a dimerization-stabilizing RAF inhibitor. In summary, we provide evidence that each of the basic residues within the RKTR motif is indispensable for correct RAF function.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
