

Peptidyl-prolyl cis/trans isomerase NIMA-interacting 1 associates with insulin receptor substrate-1 and enhances insulin actions and adipogenesis
- PMID: 21454638
- PMCID: PMC3121524
- DOI: 10.1074/jbc.M110.206904
Peptidyl-prolyl cis/trans isomerase NIMA-interacting 1 associates with insulin receptor substrate-1 and enhances insulin actions and adipogenesis
Abstract
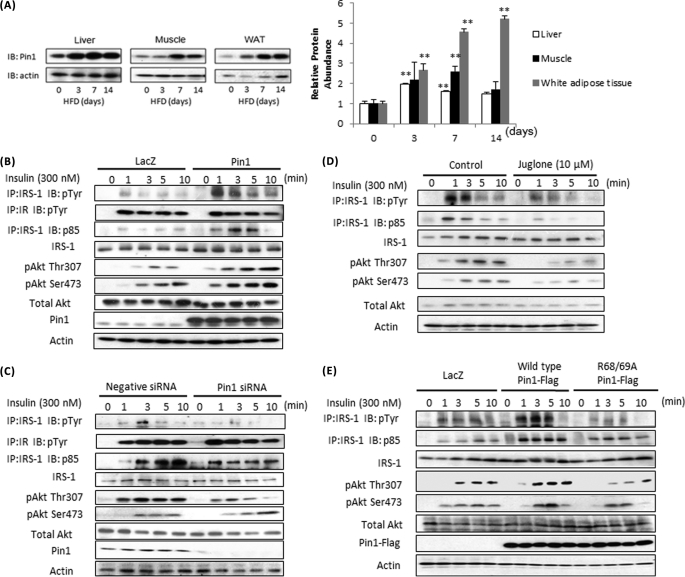
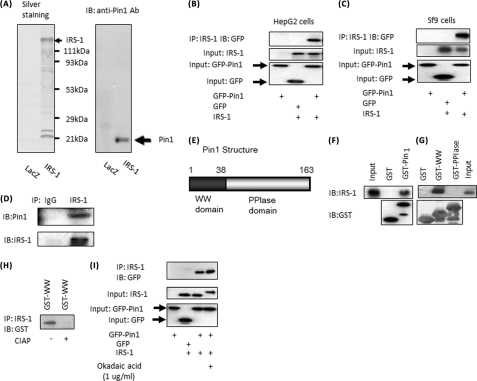
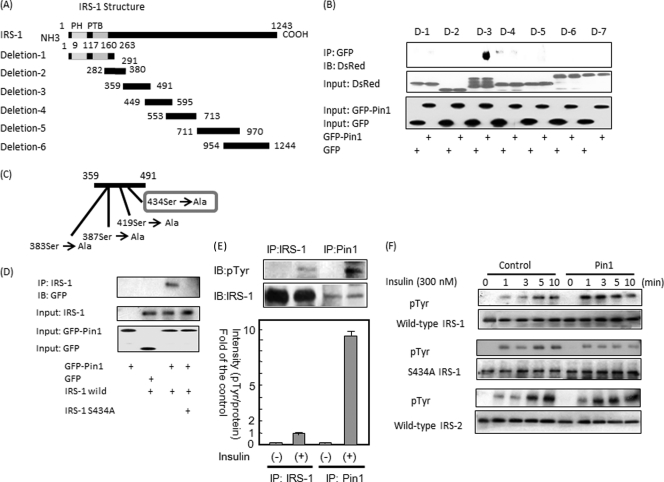
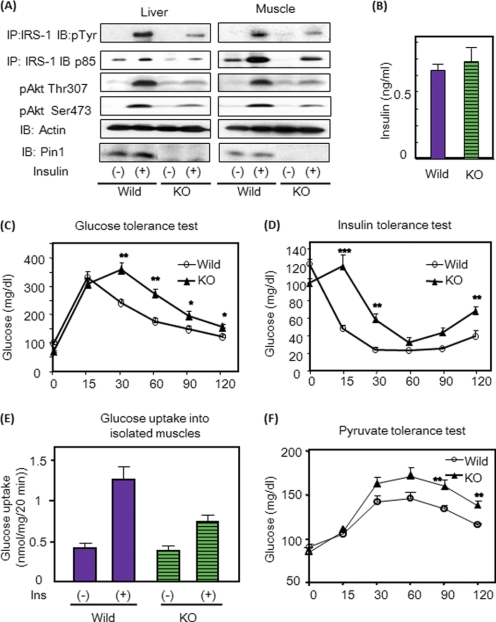
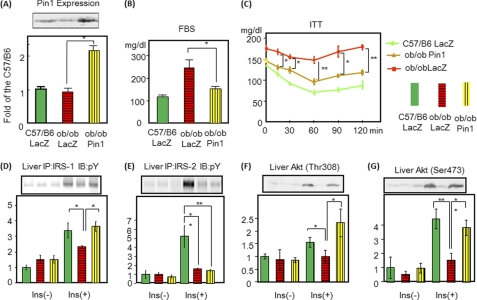
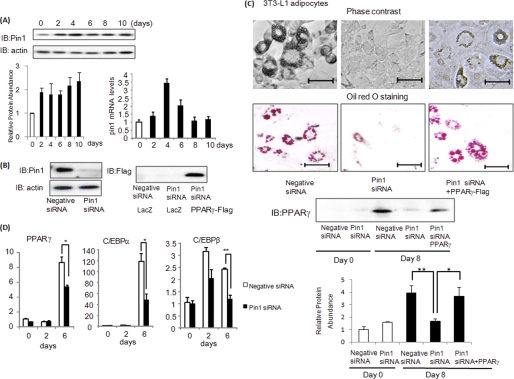
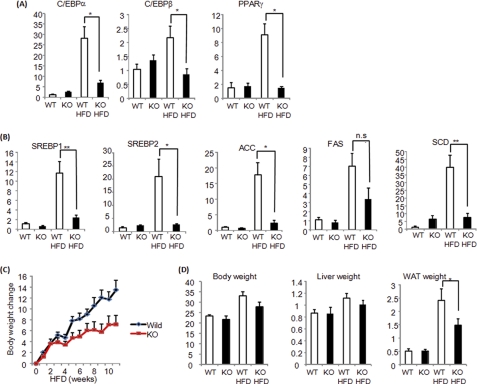
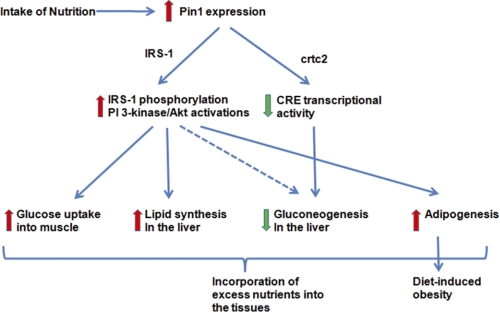
Peptidyl-prolyl cis/trans isomerase NIMA-interacting 1 (Pin1) is a unique enzyme that associates with the pSer/Thr-Pro motif and catalyzes cis-trans isomerization. We identified Pin1 in the immunoprecipitates of overexpressed IRS-1 with myc and FLAG tags in mouse livers and confirmed the association between IRS-1 and Pin1 by not only overexpression experiments but also endogenously in the mouse liver. The analysis using deletion- and point-mutated Pin1 and IRS-1 constructs revealed the WW domain located in the N terminus of Pin1 and Ser-434 in the SAIN (Shc and IRS-1 NPXY binding) domain of IRS-1 to be involved in their association. Subsequently, we investigated the role of Pin1 in IRS-1 mediation of insulin signaling. The overexpression of Pin1 in HepG2 cells markedly enhanced insulin-induced IRS-1 phosphorylation and its downstream events: phosphatidylinositol 3-kinase binding with IRS-1 and Akt phosphorylation. In contrast, the treatment of HepG2 cells with Pin1 siRNA or the Pin1 inhibitor Juglone suppressed these events. In good agreement with these in vitro data, Pin1 knock-out mice exhibited impaired insulin signaling with glucose intolerance, whereas adenoviral gene transfer of Pin1 into the ob/ob mouse liver mostly normalized insulin signaling and restored glucose tolerance. In addition, it was also demonstrated that Pin1 plays a critical role in adipose differentiation, making Pin1 knock-out mice resistant to diet-induced obesity. Importantly, Pin1 expression was shown to be up-regulated in accordance with nutrient conditions such as food intake or a high-fat diet. Taken together, these observations indicate that Pin1 binds to IRS-1 and thereby markedly enhances insulin action, essential for adipogenesis.
Figures
References
-
- Fischer G., Wittmann-Liebold B., Lang K., Kiefhaber T., Schmid F. X. (1989) Nature 337, 476–478 - PubMed
-
- Liu J., Farmer J. D., Jr., Lane W. S., Friedman J., Weissman I., Schreiber S. L. (1991) Cell 66, 807–815 - PubMed
-
- Choi J., Chen J., Schreiber S. L., Clardy J. (1996) Science 273, 239–242 - PubMed
-
- Loewith R., Jacinto E., Wullschleger S., Lorberg A., Crespo J. L., Bonenfant D., Oppliger W., Jenoe P., Hall M. N. (2002) Mol. Cell 10, 457–468 - PubMed
-
- Schreiber S. L. (1991) Science 251, 283–287 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
