

doi: 10.1104/pp.111.173708.
Epub 2011 Mar 31.
One protoplast is not the other!
Affiliations
- PMID: 21454800
- PMCID: PMC3177251
- DOI: 10.1104/pp.111.173708
Item in Clipboard
One protoplast is not the other!
Plant Physiol.
2011 Jun.
No abstract available
Figures
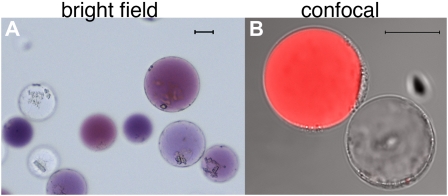
Protoplasts freshly isolated from petals of the petunia hybrid M1XV30. A, Bright-field image. The two cell types are recognizable by the presence of anthocyanins in the central vacuole of epidermal cells and their absence in the mesophyll cells. B, Confocal image of the same protoplast preparation as in A. The red fluorescence, due to anthocyanin autofluorescence, allows the recognition of the two cell types (both visible in transmitted light) during confocal analysis. The size bar equals 20 μm. Protoplast isolation: Petals or leaves from greenhouse-grown plants were sterilized in 5% hypochlorite solution (for 30 s then rinsed in sterile water) and perforated using a needle bed (a “kenzan” for Japanese ikebana), prior overnight digestion in TEX buffer (B5 salts, 500 mg/L MES, 750 mg/L CaCl2 [2*H2O] 250 mg/L NH4NO3, and 0.4 m Suc [13.7%], pH 5.7), plus 0.2% Macerozyme R10 and 0.4% Cellulase R10 (Yakult). The digested material was filtered through a 150-SIGMA mesh filter (or similar filter) and protoplast suspension was then centrifuged for 10 min at 75g at room temperature in a swing-out rotor to concentrate the protoplasts in a band floating above the medium. After 2× washing with 10 mL of TEX buffer (centrifugation at 75g for 5 min between washing steps) protoplasts were then resuspended in an appropriate volume of MMM solution (0.5 m mannitol, 15 mm MgCl2, 0.1% MES). A total of 300 μL of protoplasts was used for each transformation: 30 μg of (supercoiled) plasmid DNA was added followed by 300 μL of polyethylene glycol solution [0.4 m mannitol, 0.1 m Ca(NO3)2*H2O, 40% polyethylene glycol brought to pH 8.0 with KOH] and 2 mL of TEX. Incubation at 25°C for 2 h was followed by washing with TEX buffer, as described before, and resuspended, after centrifugation, in 2 mL of TEX buffer. We have applied this protocol to flowers of different ages (from nearly open buds to fully expanded petals) and genotypes (different genetic backgrounds and/or mutations in genes affecting pigment deposition and/or vacuolar acidification). Transformation efficiency was in all cases above 60%. Plants were grown in a greenhouse with temperature never below 19°C and never exceeding 30°C, with a cycle of a minimum of 16 h of light in all seasons (supplied with artificial light in the winter). Suboptimal or unstable plant growth conditions can make efficiency of protoplast isolation and transformation drop dramatically.
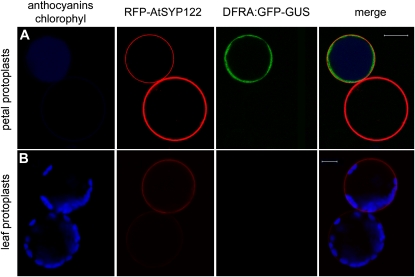
Transient expression of DFRA-GFP and 35S-RFP-AtSYP122 in petunia petal and leaf protoplasts. A, Petal protoplasts express the 35S:RFP-AtSYP122 marker of the plasma membrane (driven by the ectopic CaMV35S promoter) in both cells accumulating anthocyanins and unpigmented cells (anthocyanins in blue color to distinguish them from the RFP signal). The GFP signal (driven by the DFRa promoter) is only visible in cells accumulating anthocyanins. B, Petunia leaf protoplasts (the blue color now evidences chloroplasts) express the CaMV35S promoter driven RFP-AtSYP122, but the DFRA:GFP construct is not expressed in these cells. Images were acquired with a Zeiss confocal laser microscope (LSM Pascal). Fluorescence was detected using a 488-/543-nm dichroic beam splitter, a 505- to 530-nm band pass filter for GFP, and a 560- to 615-nm band pass filter for RFP; chlorophyll and anthocyanins epifluorescence was detected with the filter set for trimethylrhodamine isothiocyanate (>650 nm). The size bar equals 20 μm.
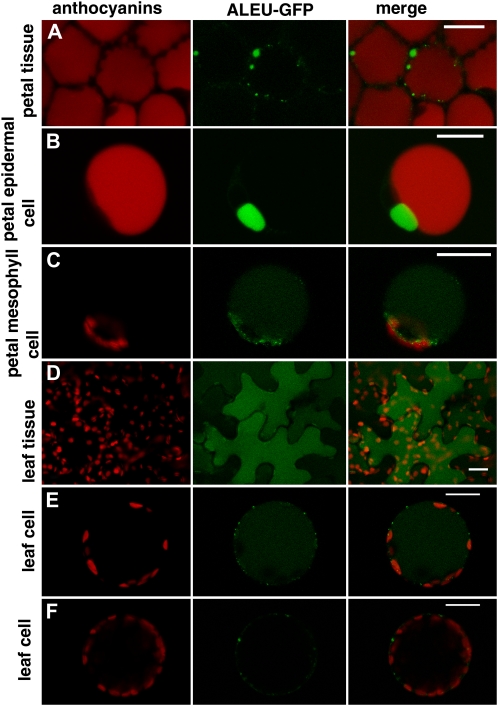
Transient expression of 35S:ALEU-GFP in petal and leaf protoplasts and in intact tissues. A, Accumulation of ALEU:GFP in petals of petunia flowers 24 h after agroinfiltration. Red autofluorescence of anthocyanins is visible in all cells and the GFP signal accumulates in small compartments independent from the central vacuole. B, Petal epidermal protoplast (recognizable from the presence of red fluorescent anthocyanins in the central vacuole) that accumulate ALEU-GFP in a small compartment independent from the central vacuole. C, Petal mesophyll protoplast (no anthocyanins in the central vacuole) accumulating ALEU-GFP in the central vacuole and in (prevacuolar) small compartments. D, Accumulation of ALEU-GFP in leaf epidermal cells 24 h after infiltration. The GFP signal is present in the central vacuole. E, Leaf protoplast (probably originating from the leaf epidermis) accumulating ALEU-GFP in the central vacuole. F, A leaf protoplast (probably originating from mesophyll) showing a different pattern of accumulation of ALEU-GFP. The size bar equals 20 μm.
References
-
- Chen S, Tao L, Zeng L, Vega-Sanchez ME, Umemura K, Wang GL. (2006) A highly efficient transient protoplast system for analyzing defence gene expression and protein-protein interactions in rice. Mol Plant Pathol 7: 417–427 - PubMed
-
- Fluckiger R, De Caroli M, Piro G, Dalessandro G, Neuhaus JM, Di Sansebastiano GP. (2003) Vacuolar system distribution in Arabidopsis tissues, visualized using GFP fusion proteins. J Exp Bot 54: 1577–1584 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
