

Motor functions of the superior colliculus
- PMID: 21456962
- PMCID: PMC3641825
- DOI: 10.1146/annurev-neuro-061010-113728
Motor functions of the superior colliculus
Abstract
The mammalian superior colliculus (SC) and its nonmammalian homolog, the optic tectum, constitute a major node in processing sensory information, incorporating cognitive factors, and issuing motor commands. The resulting action-to orient toward or away from a stimulus-can be accomplished as an integrated movement across oculomotor, cephalomotor, and skeletomotor effectors. The SC also participates in preserving fixation during intersaccadic intervals. This review highlights the repertoire of movements attributed to SC function and analyzes the significance of results obtained from causality-based experiments (microstimulation and inactivation). The mechanisms potentially used to decode the population activity in the SC into an appropriate movement command are also discussed.
Figures
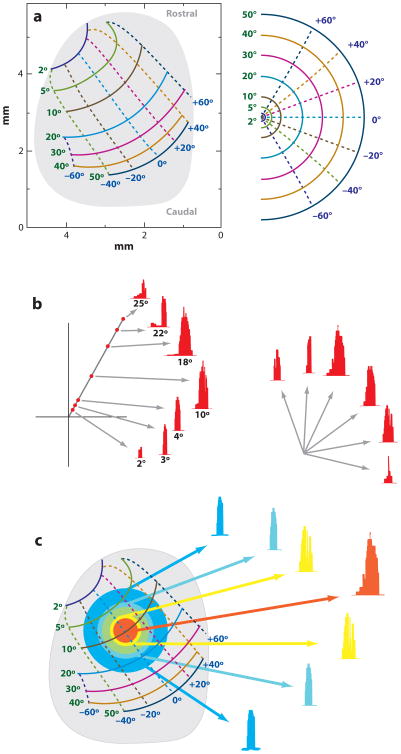
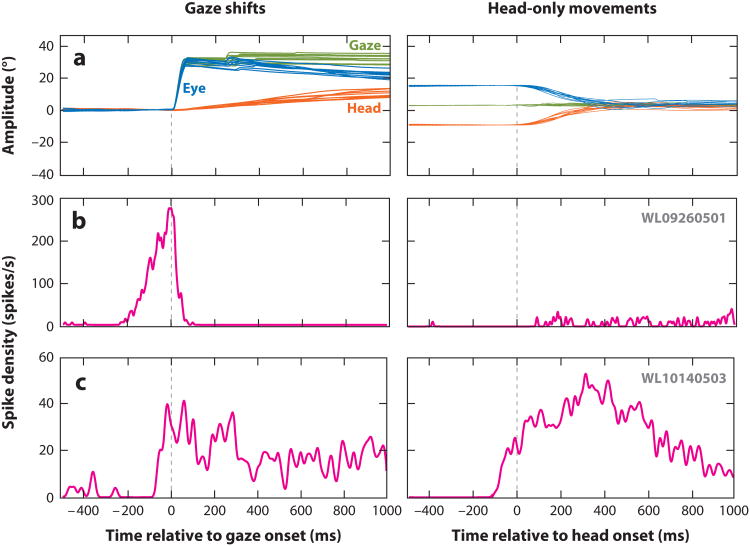
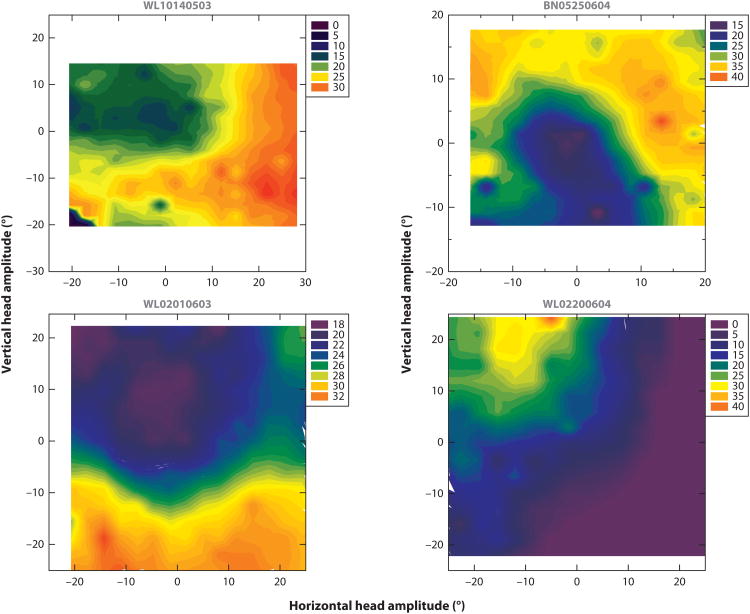
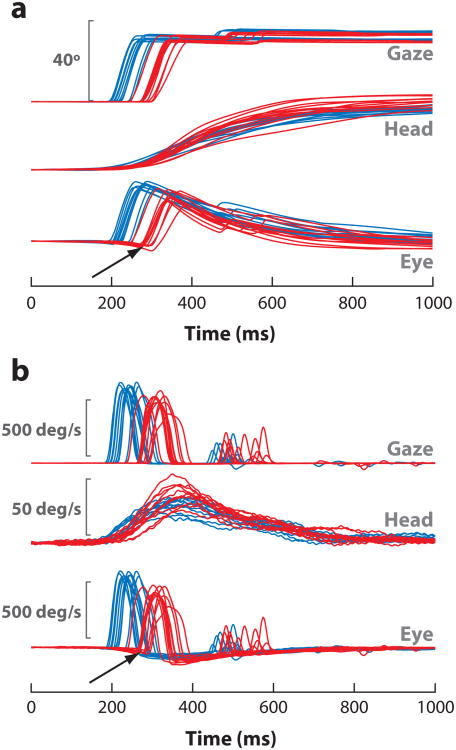
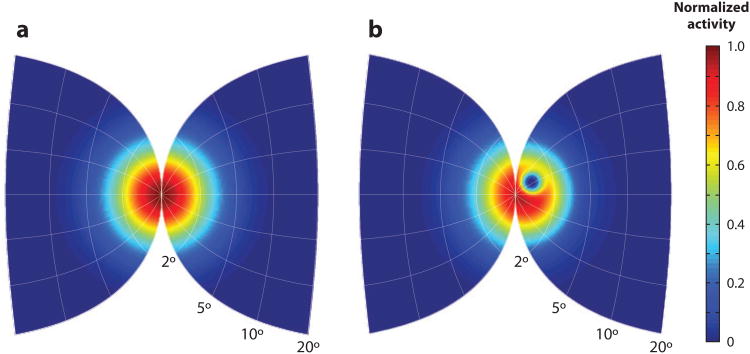
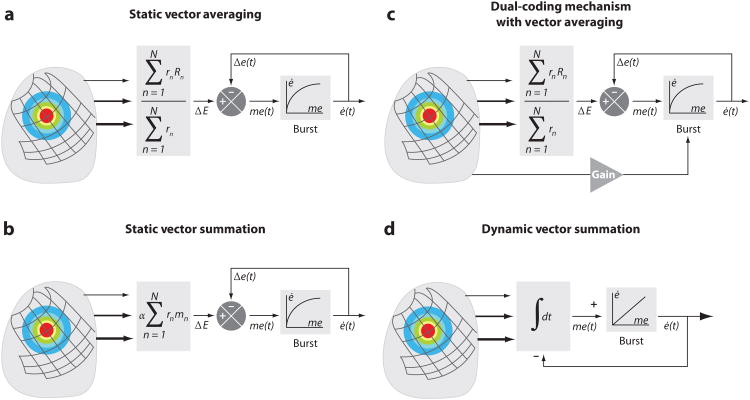
References
-
- Aizawa H, Wurtz RH. Reversible inactivation of monkey superior colliculus. I. Curvature of saccadic trajectory. J Neurophysiol. 1998;79:2082–96. - PubMed
-
- Anastasopoulos D, Ziavra N, Hollands M, Bronstein A. Gaze displacement and inter-segmental coordination during large whole body voluntary rotations. Exp Brain Res. 2009;193:323–36. - PubMed
-
- Anderson RW, Keller EL, Gandhi NJ, Das S. Two-dimensional saccade-related population activity in superior colliculus in monkey. J Neurophysiol. 1998;80:798–817. - PubMed
-
- Arai K, Keller EL, Edelman JA. Two-dimensional neural network model of the primate saccadic system. Neural Netw. 1994;7:1115–35.
-
- Badler JB, Keller EL. Decoding of a motor command vector from distributed activity in superior colliculus. Biol Cybern. 2002;86:179–89. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
