

A mutation in C2orf64 causes impaired cytochrome c oxidase assembly and mitochondrial cardiomyopathy
- PMID: 21457908
- PMCID: PMC3071910
- DOI: 10.1016/j.ajhg.2011.03.002
A mutation in C2orf64 causes impaired cytochrome c oxidase assembly and mitochondrial cardiomyopathy
Abstract
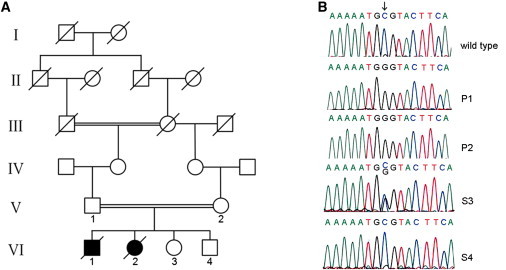
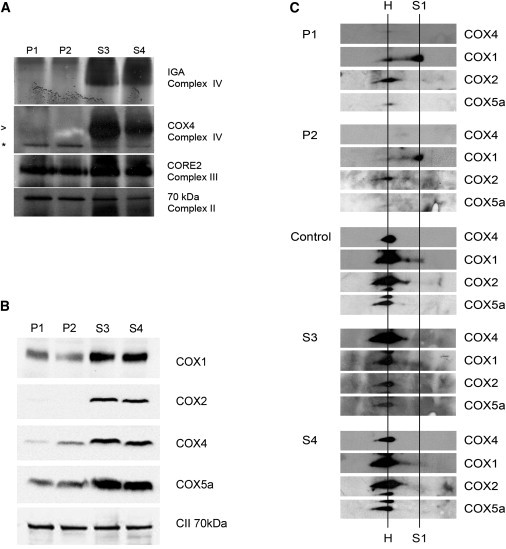
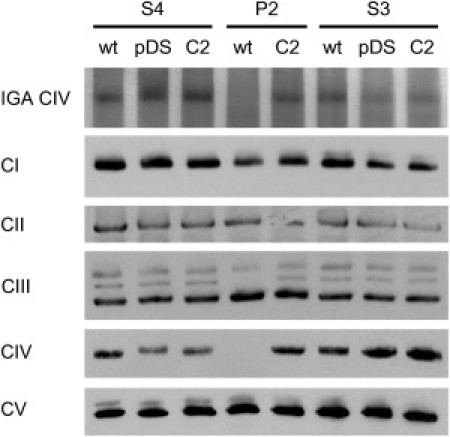
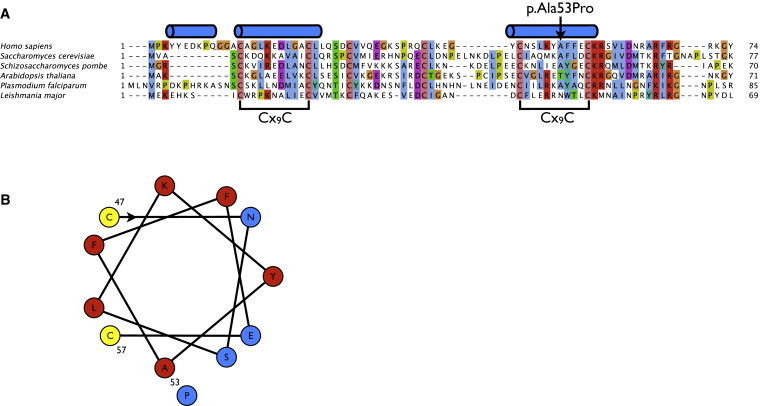
The assembly of mitochondrial respiratory chain complex IV (cytochrome c oxidase) involves the coordinated action of several assembly chaperones. In Saccharomyces cerevisiae, at least 30 different assembly chaperones have been identified. To date, pathogenic mutations leading to a mitochondrial disorder have been identified in only seven of the corresponding human genes. One of the genes for which the relevance to human pathology is unknown is C2orf64, an ortholog of the S. cerevisiae gene PET191. This gene has previously been shown to be a complex IV assembly factor in yeast, although its exact role is still unknown. Previous research in a large cohort of complex IV deficient patients did not support an etiological role of C2orf64 in complex IV deficiency. In this report, a homozygous mutation in C2orf64 is described in two siblings affected by fatal neonatal cardiomyopathy. Pathogenicity of the mutation is supported by the results of a complementation experiment, showing that complex IV activity can be fully restored by retroviral transduction of wild-type C2orf64 in patient-derived fibroblasts. Detailed analysis of complex IV assembly intermediates in patient fibroblasts by 2D-BN PAGE revealed the accumulation of a small assembly intermediate containing subunit COX1 but not the COX2, COX4, or COX5b subunits, indicating that C2orf64 is involved in an early step of the complex IV assembly process. The results of this study demonstrate that C2orf64 is essential for human complex IV assembly and that C2orf64 mutational analysis should be considered for complex IV deficient patients, in particular those with hypertrophic cardiomyopathy.
Copyright © 2011 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
A mutation in the FAM36A gene, the human ortholog of COX20, impairs cytochrome c oxidase assembly and is associated with ataxia and muscle hypotonia.Hum Mol Genet. 2013 Feb 15;22(4):656-67. doi: 10.1093/hmg/dds473. Epub 2012 Nov 2. Hum Mol Genet. 2013. PMID: 23125284
-
Whole-exome sequencing identifies a mutation in the mitochondrial ribosome protein MRPL44 to underlie mitochondrial infantile cardiomyopathy.J Med Genet. 2013 Mar;50(3):151-9. doi: 10.1136/jmedgenet-2012-101375. Epub 2013 Jan 12. J Med Genet. 2013. PMID: 23315540
-
Mutations in NDUFAF3 (C3ORF60), encoding an NDUFAF4 (C6ORF66)-interacting complex I assembly protein, cause fatal neonatal mitochondrial disease.Am J Hum Genet. 2009 Jun;84(6):718-27. doi: 10.1016/j.ajhg.2009.04.020. Epub 2009 May 21. Am J Hum Genet. 2009. PMID: 19463981 Free PMC article.
-
A novel mutation in the dihydrolipoamide dehydrogenase E3 subunit gene (DLD) resulting in an atypical form of alpha-ketoglutarate dehydrogenase deficiency.Hum Mutat. 2005 Mar;25(3):323-4. doi: 10.1002/humu.9319. Hum Mutat. 2005. PMID: 15712224 Review.
-
Cytochrome c oxidase deficiency.Am J Med Genet. 2001 Spring;106(1):46-52. doi: 10.1002/ajmg.1378. Am J Med Genet. 2001. PMID: 11579424 Review.
Cited by
-
Mutated PET117 causes complex IV deficiency and is associated with neurodevelopmental regression and medulla oblongata lesions.Hum Genet. 2017 Jun;136(6):759-769. doi: 10.1007/s00439-017-1794-7. Epub 2017 Apr 6. Hum Genet. 2017. PMID: 28386624 Free PMC article.
-
Mitochondrial cytochrome c oxidase deficiency.Clin Sci (Lond). 2016 Mar;130(6):393-407. doi: 10.1042/CS20150707. Clin Sci (Lond). 2016. PMID: 26846578 Free PMC article. Review.
-
Cysteine residues in mitochondrial intermembrane space proteins: more than just import.Br J Pharmacol. 2019 Feb;176(4):514-531. doi: 10.1111/bph.14480. Epub 2018 Sep 28. Br J Pharmacol. 2019. PMID: 30129023 Free PMC article. Review.
-
Building the CuA site of cytochrome c oxidase: A complicated, redox-dependent process driven by a surprisingly large complement of accessory proteins.J Biol Chem. 2018 Mar 30;293(13):4644-4652. doi: 10.1074/jbc.R117.816132. Epub 2017 Sep 29. J Biol Chem. 2018. PMID: 28972150 Free PMC article. Review.
-
The clinical maze of mitochondrial neurology.Nat Rev Neurol. 2013 Aug;9(8):429-44. doi: 10.1038/nrneurol.2013.126. Epub 2013 Jul 9. Nat Rev Neurol. 2013. PMID: 23835535 Free PMC article. Review.
References
-
- Shoubridge E.A. Cytochrome c oxidase deficiency. Am. J. Med. Genet. 2001;106:46–52. - PubMed
-
- Massa V., Fernandez-Vizarra E., Alshahwan S., Bakhsh E., Goffrini P., Ferrero I., Mereghetti P., D'Adamo P., Gasparini P., Zeviani M. Severe infantile encephalomyopathy caused by a mutation in COX6B1, a nucleus-encoded subunit of cytochrome c oxidase. Am. J. Hum. Genet. 2008;82:1281–1289. - PMC - PubMed
-
- Weraarpachai W., Antonicka H., Sasarman F., Seeger J., Schrank B., Kolesar J.E., Lochmüller H., Chevrette M., Kaufman B.A., Horvath R., Shoubridge E.A. Mutation in TACO1, encoding a translational activator of COX I, results in cytochrome c oxidase deficiency and late-onset Leigh syndrome. Nat. Genet. 2009;41:833–837. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
