

Bone marrow mesenchymal stem and progenitor cells induce monocyte emigration in response to circulating toll-like receptor ligands
- PMID: 21458307
- PMCID: PMC3081416
- DOI: 10.1016/j.immuni.2011.02.016
Bone marrow mesenchymal stem and progenitor cells induce monocyte emigration in response to circulating toll-like receptor ligands
Abstract
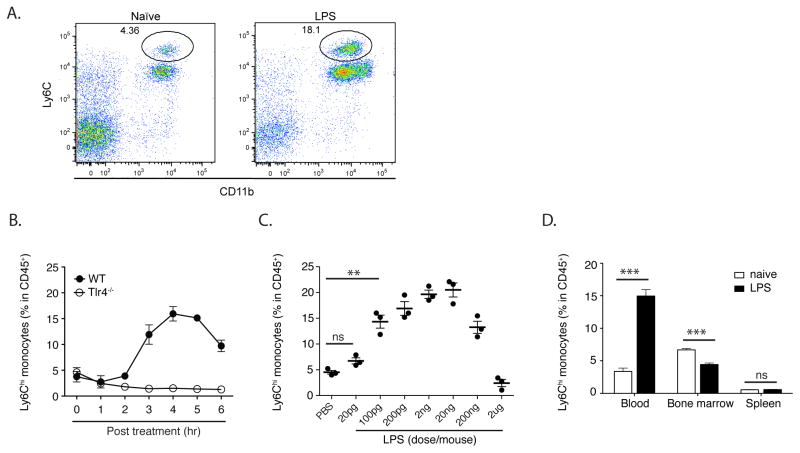
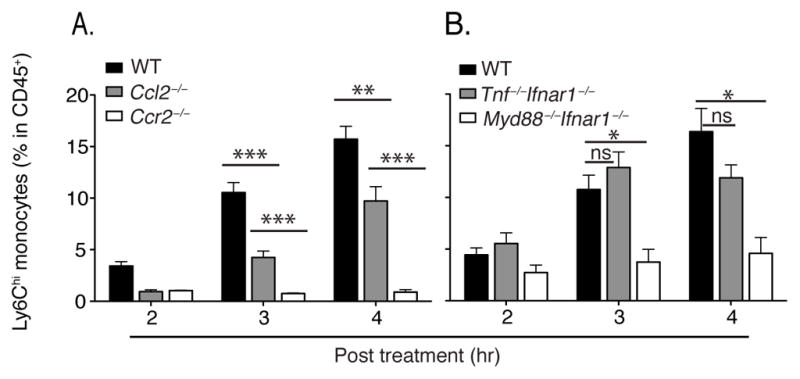
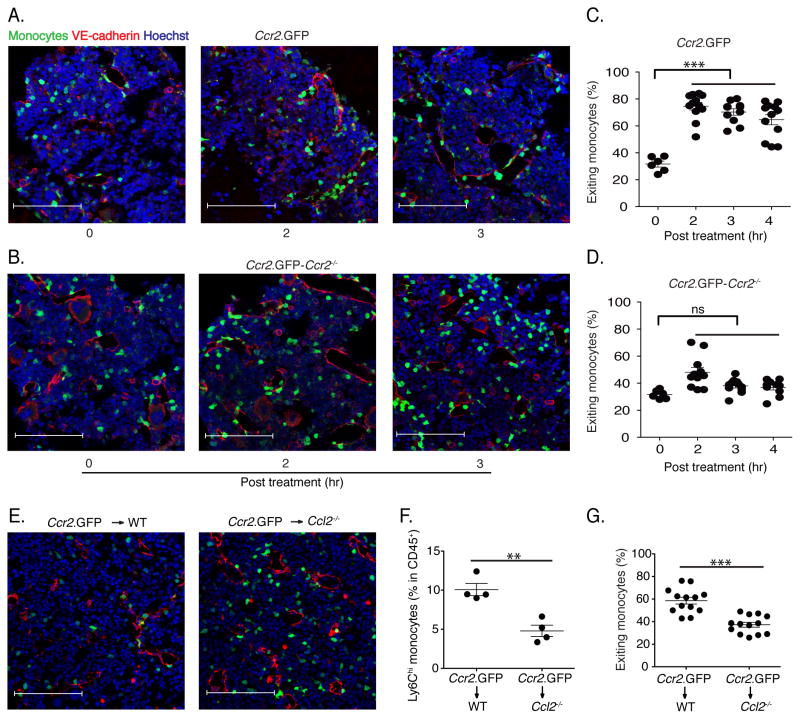
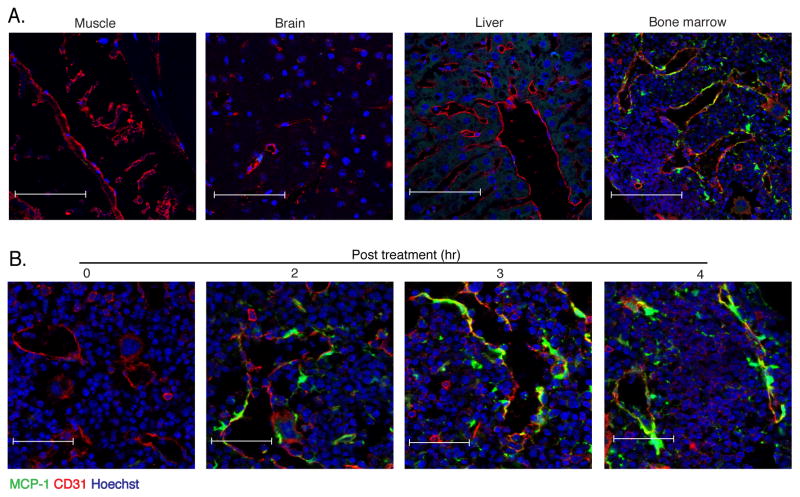
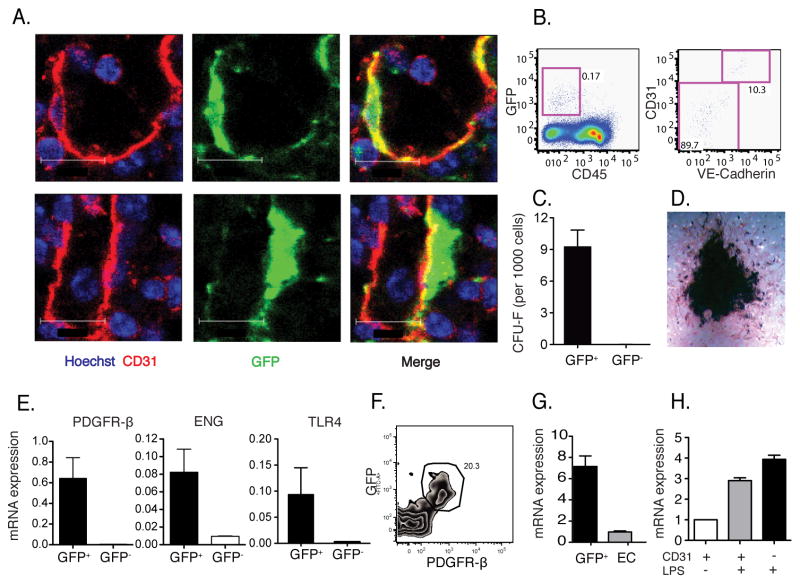
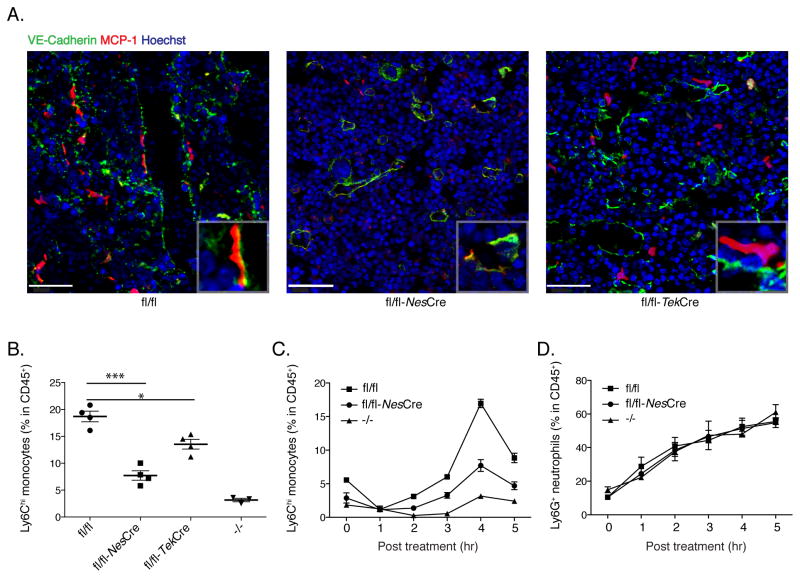
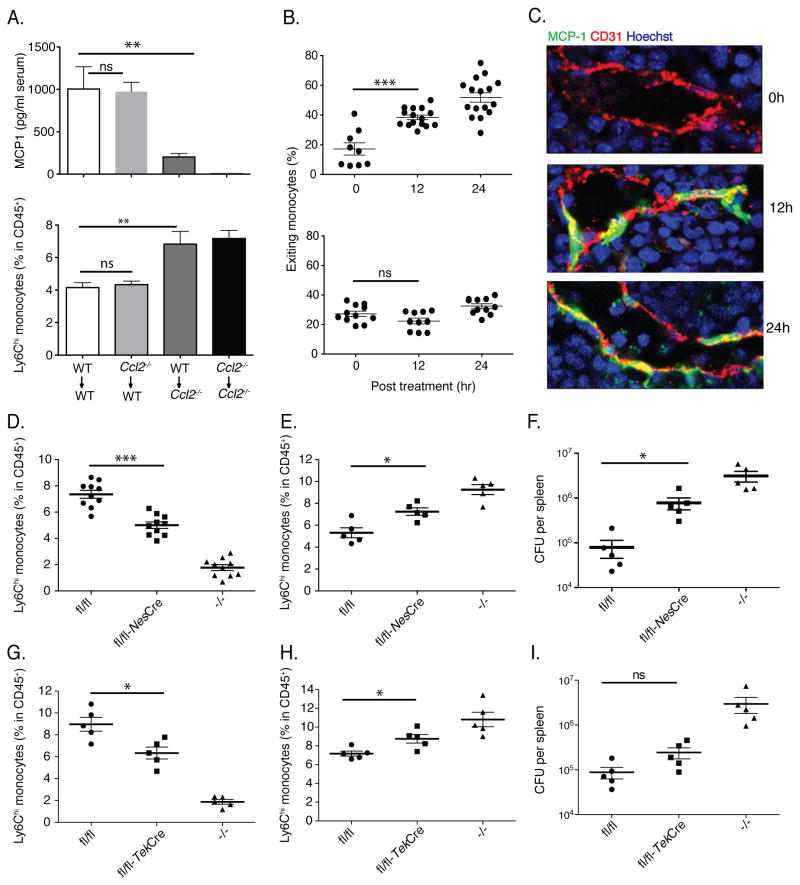
Inflammatory (Ly6C(hi) CCR2+) monocytes provide defense against infections but also contribute to autoimmune diseases and atherosclerosis. Monocytes originate from bone marrow and their entry into the bloodstream requires stimulation of CCR2 chemokine receptor by monocyte chemotactic protein-1 (MCP1). How monocyte emigration from bone marrow is triggered by remote infections remains unclear. We demonstrated that low concentrations of Toll-like receptor (TLR) ligands in the bloodstream drive CCR2-dependent emigration of monocytes from bone marrow. Bone marrow mesenchymal stem cells (MSCs) and their progeny, including CXC chemokine ligand (CXCL)12-abundant reticular (CAR) cells, rapidly expressed MCP1 in response to circulating TLR ligands or bacterial infection and induced monocyte trafficking into the bloodstream. Targeted deletion of MCP1 from MSCs impaired monocyte emigration from bone marrow. Our findings suggest that bone marrow MSCs and CAR cells respond to circulating microbial molecules and regulate bloodstream monocyte frequencies by secreting MCP1 in proximity to bone marrow vascular sinuses.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Emergency evacuation! Hematopoietic niches induce cell exit in infection.Immunity. 2011 Apr 22;34(4):463-5. doi: 10.1016/j.immuni.2011.04.009. Immunity. 2011. PMID: 21511181
-
Monocytes: nudged out of the niche.Nat Rev Immunol. 2011 Jun;11(6):368-9. doi: 10.1038/nri2999. Nat Rev Immunol. 2011. PMID: 21610733 No abstract available.
References
-
- Boring L, Gosling J, Cleary M, Charo IF. Decreased lesion formation in CCR2−/− mice reveals a role for chemokines in the initiation of atherosclerosis. Nature. 1998;394:894–897. - PubMed
-
- Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, Kazzaz Z, Bornstein E, Lambotte O, Altmann D, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006;12:1365–1371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 5P01CA023766-31/CA/NCI NIH HHS/United States
- P01 CA023766/CA/NCI NIH HHS/United States
- R01 AI080619/AI/NIAID NIH HHS/United States
- R01 HL069438/HL/NHLBI NIH HHS/United States
- R01 HL097819/HL/NHLBI NIH HHS/United States
- 5R37AI039031/AI/NIAID NIH HHS/United States
- R37 AI039031/AI/NIAID NIH HHS/United States
- R01DK056638/DK/NIDDK NIH HHS/United States
- K08AI071998/AI/NIAID NIH HHS/United States
- R01 DK056638/DK/NIDDK NIH HHS/United States
- K08 AI071998/AI/NIAID NIH HHS/United States
- R01HL097819/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
