

The metabolic bioactivation of caffeic acid phenethyl ester (CAPE) mediated by tyrosinase selectively inhibits glutathione S-transferase
- PMID: 21458432
- PMCID: PMC3706206
- DOI: 10.1016/j.cbi.2011.03.015
The metabolic bioactivation of caffeic acid phenethyl ester (CAPE) mediated by tyrosinase selectively inhibits glutathione S-transferase
Abstract
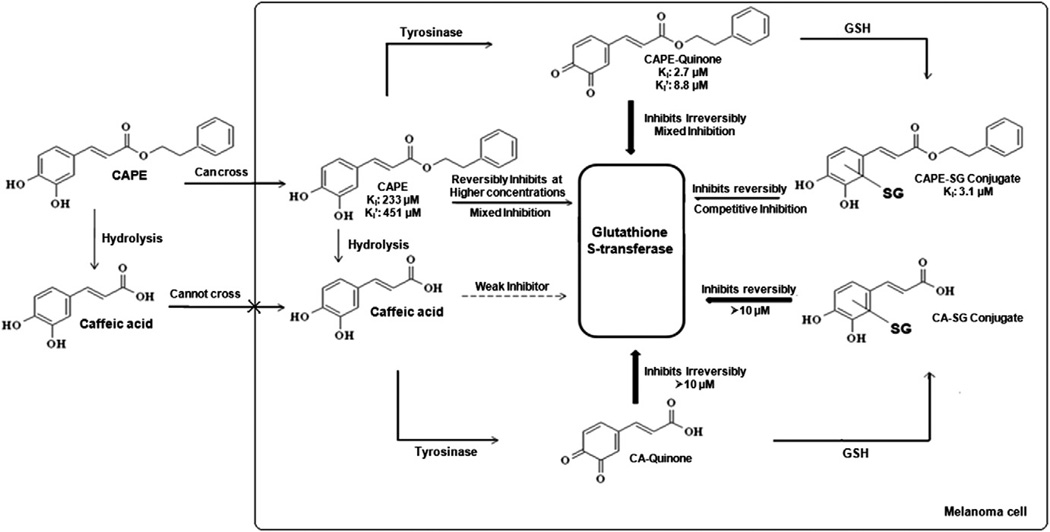
Glutathione S-transferase (GST) and multidrug resistance-associated proteins (MRPs) play major roles in drug resistance in melanoma. In this study, we investigated caffeic acid phenethyl ester (CAPE) as a selective GST inhibitor in the presence of tyrosinase, which is abundant in melanoma cells. Tyrosinase bioactivates CAPE to an o-quinone, which reacts with glutathione to form CAPE-SG conjugate. Our findings indicate that 90% CAPE was metabolized by tyrosinase after a 60-min incubation. LC-MS/MS analyses identified a CAPE-SG conjugate as a major metabolite. In the presence of tyrosinase, CAPE (10-25μM) showed 70-84% GST inhibition; whereas in the absence of tyrosinase, CAPE did not inhibit GST. CAPE-SG conjugate and CAPE-quinone (25μM) demonstrated ⩾85% GST inhibition via reversible and irreversible mechanisms, respectively. Comparing with CDNB and GSH, the non-substrate CAPE acted as a weak, reversible GST inhibitor at concentrations >50μM. Furthermore, MK-571, a selective MRP inhibitor, and probenecid, a non-selective MRP inhibitor, decrease the IC(50) of CAPE (15μM) by 13% and 21%, apoptotic cell death by 3% and 13%, and mitochondrial membrane potential in human SK-MEL-28 melanoma cells by 10% and 56%, respectively. Moreover, computational docking analyses suggest that CAPE binds to the GST catalytic active site. Caffeic acid, a hydrolyzed product of CAPE, showed a similar GST inhibition in the presence of tyrosinase. Although, as controls, 4-hydroxyanisole and L-tyrosine were metabolized by tyrosinase to form quinones and glutathione conjugates, they exhibited no GST inhibition in the absence and presence of tyrosinase. In conclusion, both CAPE and caffeic acid selectively inhibited GST in the presence of tyrosinase. Our results suggest that intracellularly formed quinones and glutathione conjugates of caffeic acid and CAPE may play major roles in the selective inhibition of GST in SK-MEL-28 melanoma cells. Moreover, the inhibition of MRP enhances CAPE-induced toxicity in the SK-MEL-28 melanoma cells.
Copyright © 2011 Elsevier Ireland Ltd. All rights reserved.
Conflict of interest statement
The authors state no conflict of interest.
Figures








References
-
- Kudugunti SK, Vad NM, Ekogbo E, Moridani MY. Efficacy of caffeic acid phenethyl ester (CAPE) in skin B16-F0 melanoma tumor bearing C57BL/6 mice. Invest. New Drugs. 2009;29(1):52–62. - PubMed
-
- Banerjee BD, Seth V, Bhattacharya A, Pasha ST, Chakraborty AK. Biochemical effects of some pesticides on lipid peroxidation and free-radical scavengers. Toxicol. Lett. 1999;107(1–3):33–47. - PubMed
-
- Sau A, Pellizzari Tregno F, Valentino F, Federici G, Caccuri AM. Glutathione transferases and development of new principles to overcome drug resistance. Arch. Biochem. Biophys. 2010;500(2):116–122. - PubMed
-
- Mena S, Ortega A, Estrela JM. Oxidative stress in environmental-induced carcinogenesis. Mutat. Res. 2009;674(1–2):36–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
