

The Mre11:Rad50 structure shows an ATP-dependent molecular clamp in DNA double-strand break repair
- PMID: 21458667
- PMCID: PMC3071652
- DOI: 10.1016/j.cell.2011.02.038
The Mre11:Rad50 structure shows an ATP-dependent molecular clamp in DNA double-strand break repair
Abstract
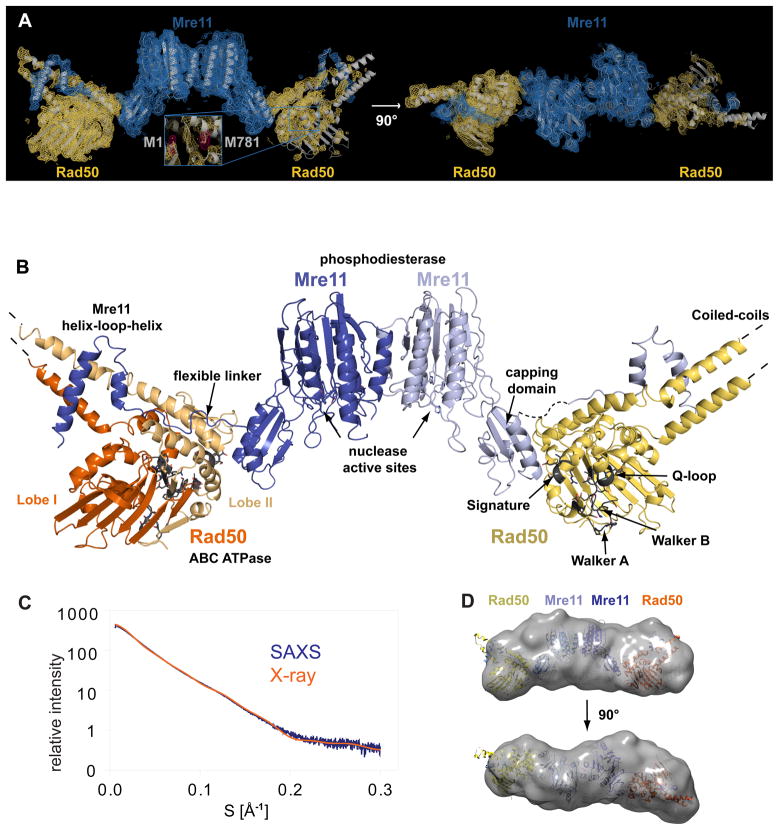
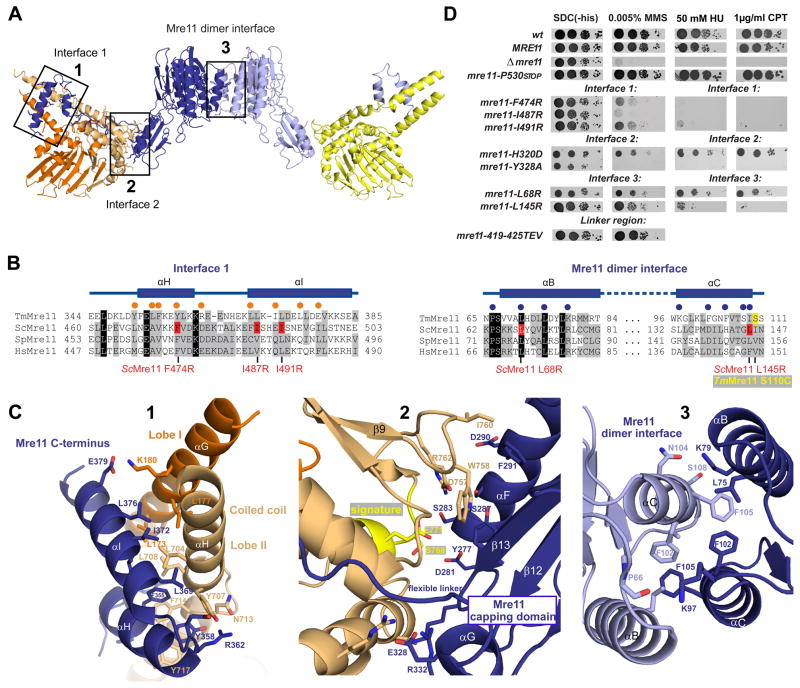
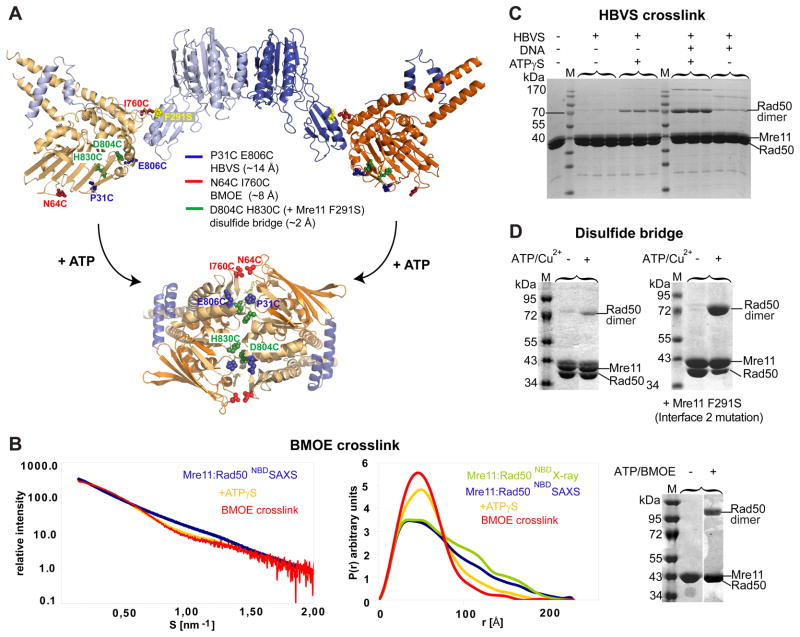
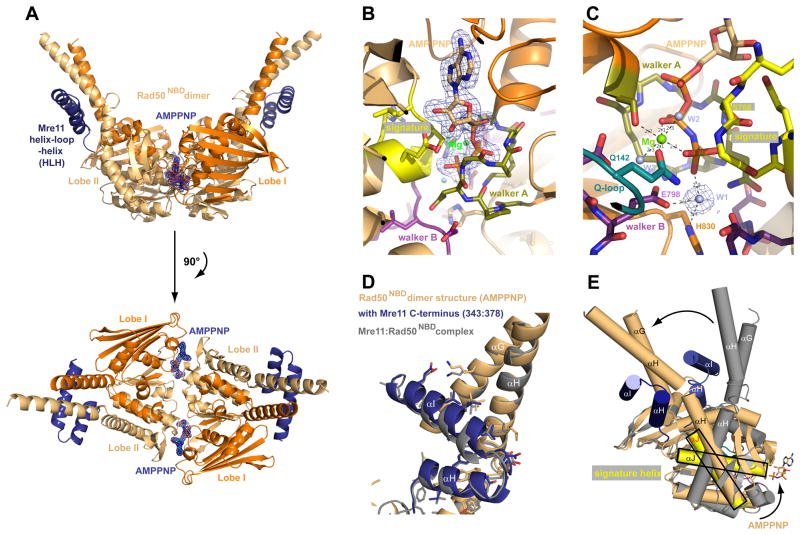
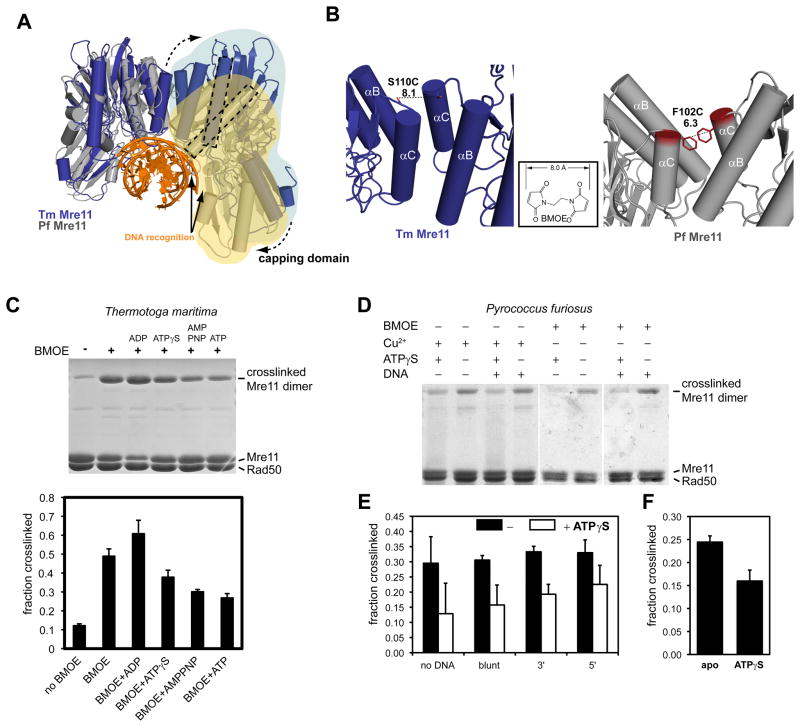
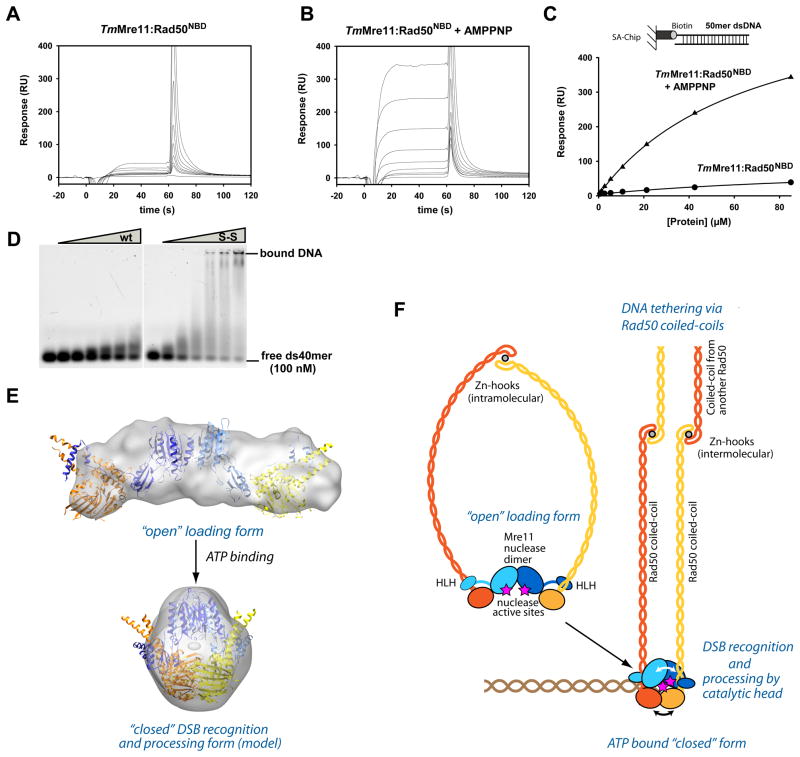
The MR (Mre11 nuclease and Rad50 ABC ATPase) complex is an evolutionarily conserved sensor for DNA double-strand breaks, highly genotoxic lesions linked to cancer development. MR can recognize and process DNA ends even if they are blocked and misfolded. To reveal its mechanism, we determined the crystal structure of the catalytic head of Thermotoga maritima MR and analyzed ATP-dependent conformational changes. MR adopts an open form with a central Mre11 nuclease dimer and two peripheral Rad50 molecules, a form suited for sensing obstructed breaks. The Mre11 C-terminal helix-loop-helix domain binds Rad50 and attaches flexibly to the nuclease domain, enabling large conformational changes. ATP binding to the two Rad50 subunits induces a rotation of the Mre11 helix-loop-helix and Rad50 coiled-coil domains, creating a clamp conformation with increased DNA-binding activity. The results suggest that MR is an ATP-controlled transient molecular clamp at DNA double-strand breaks.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
