

PARP-2 regulates SIRT1 expression and whole-body energy expenditure
- PMID: 21459329
- PMCID: PMC3108571
- DOI: 10.1016/j.cmet.2011.03.013
PARP-2 regulates SIRT1 expression and whole-body energy expenditure
Abstract
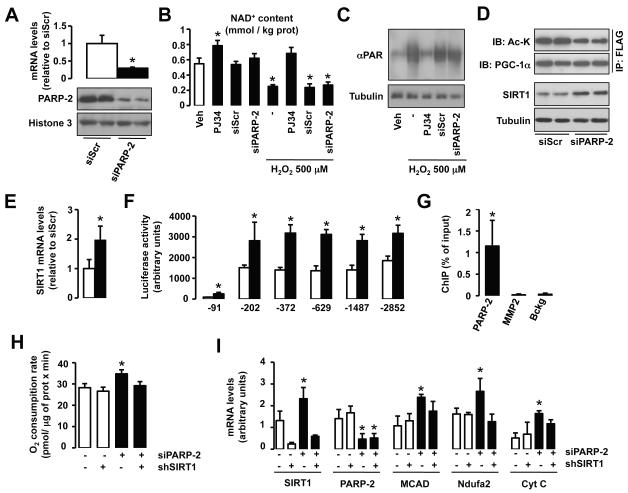
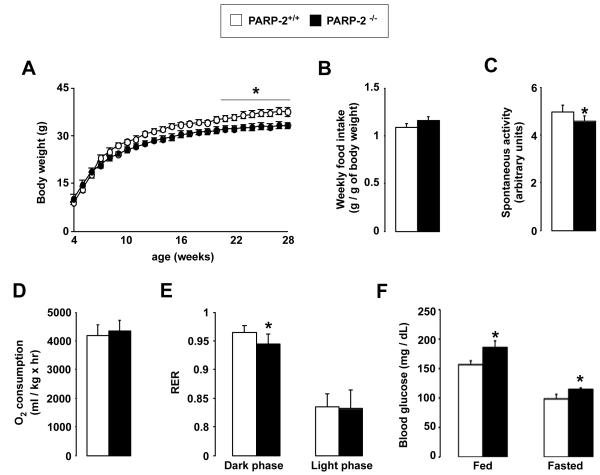
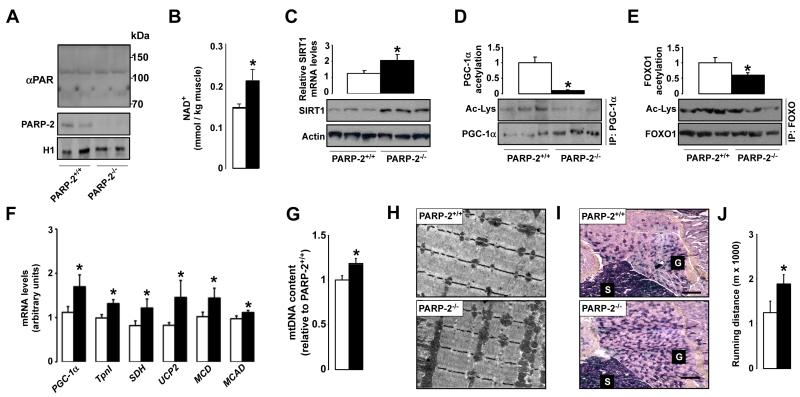
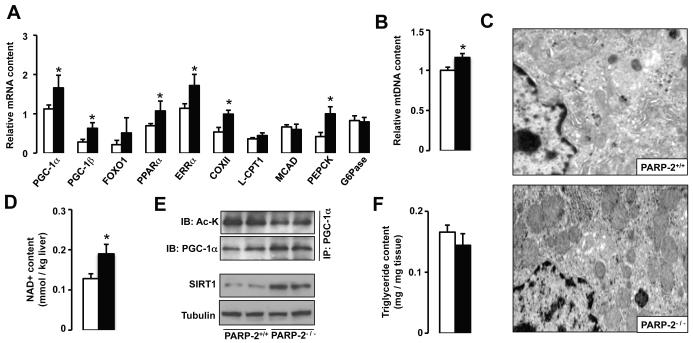
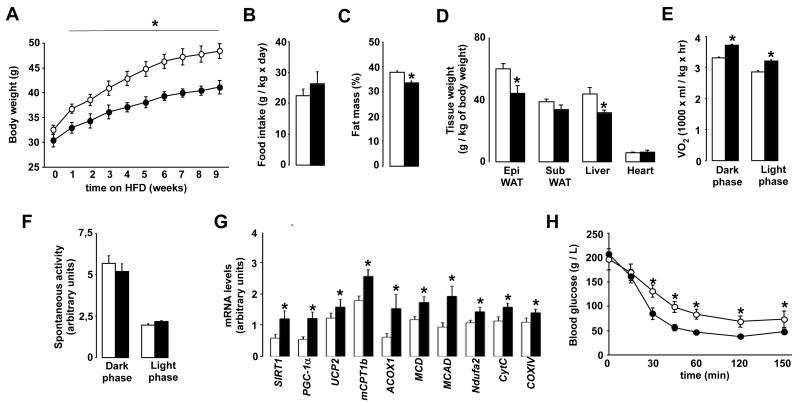
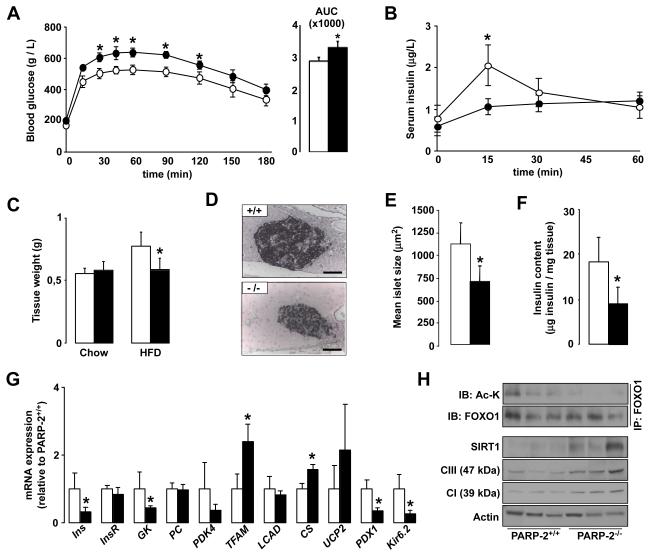
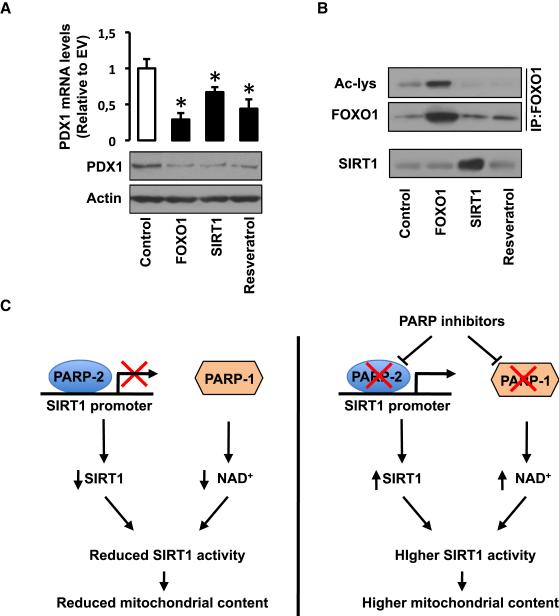
SIRT1 is a NAD(+)-dependent enzyme that affects metabolism by deacetylating key transcriptional regulators of energy expenditure. Here, we tested whether deletion of PARP-2, an alternative NAD(+)-consuming enzyme, impacts on NAD(+) bioavailability and SIRT1 activity. Our results indicate that PARP-2 deficiency increases SIRT1 activity in cultured myotubes. However, this increase was not due to changes in NAD(+) levels, but to an increase in SIRT1 expression, as PARP-2 acts as a direct negative regulator of the SIRT1 promoter. PARP-2 deletion in mice increases SIRT1 levels, promotes energy expenditure, and increases mitochondrial content. Furthermore, PARP-2(-/-) mice were protected against diet-induced obesity. Despite being insulin sensitized, PARP-2(-/-) mice were glucose intolerant due to a defective pancreatic function. Hence, while inhibition of PARP activity promotes oxidative metabolism through SIRT1 activation, the use of PARP inhibitors for metabolic purposes will require further understanding of the specific functions of different PARP family members.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A one and a two … expanding roles for poly(ADP-ribose) polymerases in metabolism.Cell Metab. 2011 Apr 6;13(4):353-355. doi: 10.1016/j.cmet.2011.03.011. Cell Metab. 2011. PMID: 21459317 Free PMC article.
References
-
- Ame JC, Rolli V, Schreiber V, Niedergang C, Apiou F, Decker P, Muller S, Hoger T, Menissier-de Murcia J, de Murcia G. PARP-2, A novel mammalian DNA damage-dependent poly(ADP-ribose) polymerase. J.Biol.Chem. 1999;274:17860–17868. - PubMed
-
- Bai P, Houten SM, Huber A, Schreiber V, Watanabe M, Kiss B, de Murcia G, Auwerx J, Menissier-de Murcia J. Poly(ADP-ribose) polymerase-2 controls adipocyte differentiation and adipose tissue function through the regulation of the activity of the retinoid × receptor/peroxisome proliferator-activated receptor-gamma heterodimer. J.Biol.Chem. 2007;282:37738–37746. - PubMed
-
- Brunet A, Sweeney LB, Sturgill JF, Chua KF, Greer PL, Lin Y, Tran H, Ross SE, Mostoslavsky R, Cohen HY, Hu LS, Cheng HL, Jedrychowski MP, Gygi SP, Sinclair DA, Alt FW, Greenberg ME. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004;303:2011–2015. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
