

The oral commensal, Streptococcus gordonii, synergizes with Tat protein to induce HIV-1 promoter activation in monocytes/macrophages
- PMID: 21459369
- PMCID: PMC3115710
- DOI: 10.1016/j.cellimm.2011.03.008
The oral commensal, Streptococcus gordonii, synergizes with Tat protein to induce HIV-1 promoter activation in monocytes/macrophages
Abstract
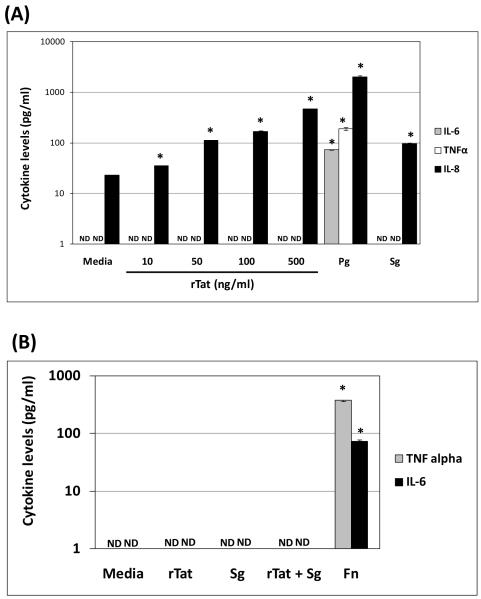
Trans-activator of transcription (Tat) is an HIV-1 protein essential for viral replication. Oral periodontopathogens (e.g. Fusobacterium nucleatum) enhance HIV-1LTR promoter activation in monocytes/macrophages in absence of Tat; however, some oral commensals fail to trigger this response. We sought to determine the effect of Tat on HIV-1LTR promoter activation induced by the representative oral commensal Streptococcus gordonii in monocytes/macrophages. S. gordonii enhanced HIV-1LTR reactivation in THP89GFP (Tat(+)), but not in BF24 (Tat(-)) cells. Interestingly, S. gordonii, but not Streptococcus sanguinis enhanced HIV-1LTR activation in the presence of recombinant Tat in BF24 cells. This response correlated with IL-8 but not TNFα or IL-6 production, and was abrogated by the NFκB inhibitor BAY 11-7082. Kinetics of NFκB-RelA activation did not explain the S. gordonii-induced HIV-1LTR activation in presence of Tat. These results suggest that S. gordonii-induced HIV-1 reactivation in monocytes/macrophages is Tat-dependent and appears to involve NFκB activation.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
HIV-1 reactivation induced by the periodontal pathogens Fusobacterium nucleatum and Porphyromonas gingivalis involves Toll-like receptor 2 [corrected] and 9 activation in monocytes/macrophages.Clin Vaccine Immunol. 2010 Sep;17(9):1417-27. doi: 10.1128/CVI.00009-10. Epub 2010 Jul 7. Clin Vaccine Immunol. 2010. PMID: 20610663 Free PMC article.
-
Supernatants from oral epithelial cells and gingival fibroblasts modulate human immunodeficiency virus type 1 promoter activation induced by periodontopathogens in monocytes/macrophages.Mol Oral Microbiol. 2010 Apr;25(2):136-49. doi: 10.1111/j.2041-1014.2009.00552.x. Mol Oral Microbiol. 2010. PMID: 20331801 Free PMC article.
-
Effects of the tat and nef gene products of human immunodeficiency virus type 1 (HIV-1) on transcription controlled by the HIV-1 long terminal repeat and on cell growth in macrophages.J Virol. 1993 Dec;67(12):6956-64. doi: 10.1128/JVI.67.12.6956-6964.1993. J Virol. 1993. PMID: 8230418 Free PMC article.
-
Tat protein induces human immunodeficiency virus type 1 (HIV-1) coreceptors and promotes infection with both macrophage-tropic and T-lymphotropic HIV-1 strains.J Virol. 1998 Nov;72(11):8952-60. doi: 10.1128/JVI.72.11.8952-8960.1998. J Virol. 1998. PMID: 9765440 Free PMC article.
-
HIV-1 Tat protein induces IL-10 production in monocytes by classical and alternative NF-kappaB pathways.Eur J Cell Biol. 2008 Dec;87(12):947-62. doi: 10.1016/j.ejcb.2008.06.005. Epub 2008 Aug 29. Eur J Cell Biol. 2008. PMID: 18760861
Cited by
-
Macrophage polarization in response to oral commensals and pathogens.Pathog Dis. 2016 Apr;74(3):ftw011. doi: 10.1093/femspd/ftw011. Epub 2016 Feb 15. Pathog Dis. 2016. PMID: 26884502 Free PMC article.
-
Interaction between endogenous bacterial flora and latent HIV infection.Clin Vaccine Immunol. 2013 Jun;20(6):773-9. doi: 10.1128/CVI.00766-12. Epub 2013 Apr 24. Clin Vaccine Immunol. 2013. PMID: 23616411 Free PMC article. Review.
References
-
- Richman DD, Margolis DM, Delaney M, Greene WC, Hazuda D, Pomerantz RJ. The challenge of finding a cure for HIV infection. Science. 2009;323:1304–1307. - PubMed
-
- Rohr O, Marban C, Aunis D, Schaeffer E. Regulation of HIV-1 gene transcription: from lymphocytes to microglial cells. J Leukoc Biol. 2003;74:736–749. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
