

doi: 10.1038/nsmb.2041.
Epub 2011 Apr 3.
Structure of catalytically competent intein caught in a redox trap with functional and evolutionary implications
Affiliations
- PMID: 21460844
- PMCID: PMC3087850
- DOI: 10.1038/nsmb.2041
Item in Clipboard
Structure of catalytically competent intein caught in a redox trap with functional and evolutionary implications
Nat Struct Mol Biol.
2011 May.
Abstract
Here we describe self-splicing proteins, called inteins, that function as redox-responsive switches in bacteria. Redox regulation was achieved by engineering a disulfide bond between the intein's catalytic cysteine and a cysteine in the flanking 'extein' sequence. This interaction was validated by an X-ray structure, which includes a transient splice junction. A natural analog of the designed system was identified in Pyrococcus abyssi, suggesting an unprecedented form of adaptive, post-translational regulation.
Figures
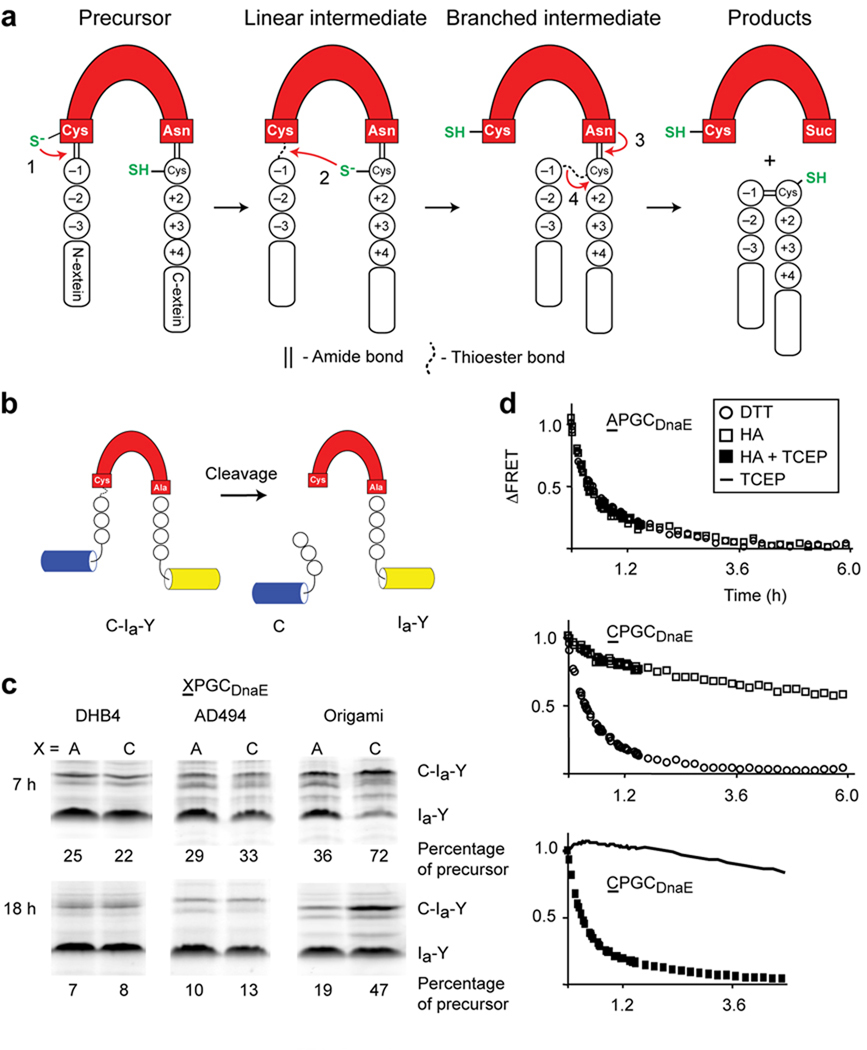
Engineered redox-responsive intein precursor in vivo and in vitro. (a) Scheme for intein splicing. Conserved terminal intein residues are indicated as is the extein C+1 residue. N-extein and C-extein residues are designated by minus and plus signs, respectively. (b). FRET-based intein reporter with the DnaE intein (red) inserted between cyan and yellow fluorescent proteins. (c). Biomimetic Cys-3 variant, CPGCDnaE, displays enhanced redox sensitivity in vivo. Activities of CPGCDnaE and APGCDnaE inteins were quantified as the percentage of precursor (C-Ia-Y) remaining, 7 h and 18 h after induction of protein expression in E. coli DHB4, AD494 and origami. Data are representative of ≥2 independent experiments. Additional bands, observed previously with unboiled samples, were excluded from the quantitation (d). Effect of redox conditions on the in vitro cleavage kinetics of APGCDnaE and CPGCDnaE. N-extein cleavage was monitored by FRET loss that occurs upon cleavage by DTT or by hydroxylamine (HA). Each data point is an average of three measurements.
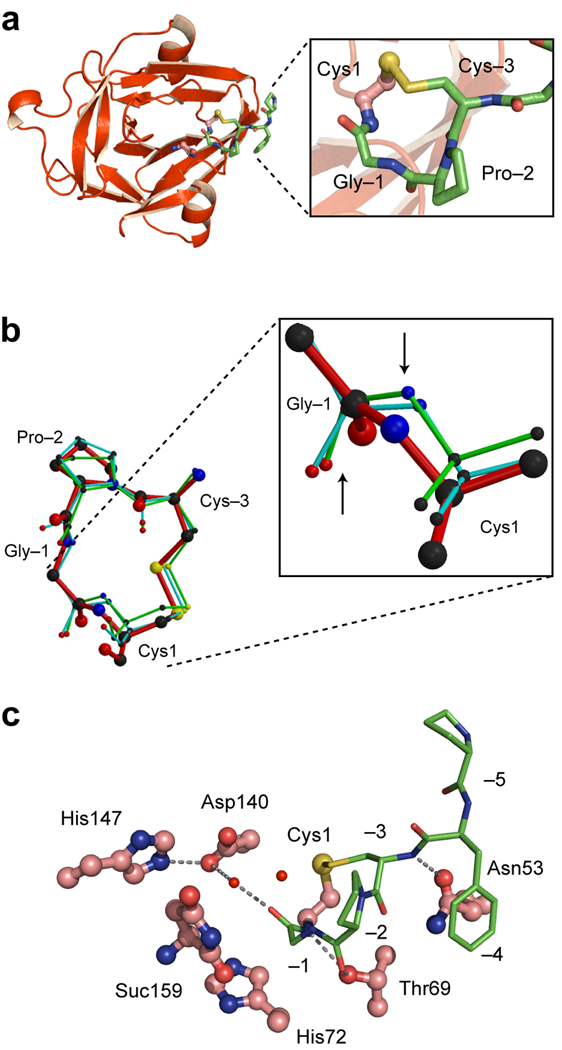
Crystal structure of the CPGCDnaE intein. (a). Ribbon diagram showing the intein (red) and the N-terminal extein (green). Magnified view of the Cys1-to-Cys-3 disulfide loop is shown in the box (b). Conformation of the CPGC disulfide loop. Comparison of the structure of the disulfide loop in the CPGCDnaE intein (red) with the two other observations of this loop in the Protein Data Bank (1JBQ, green; 1M6Y, cyan). Magnified view shows Gly-Cys peptide bonds, with an upward arrow pointing to the carbonyl oxygen atoms and a downward arrow pointing toward the amide nitrogen atoms. (c). Active-site interactions in the CPGCDnaE intein (intein, red; extein, green). Thr69 of the conserved TXXH motif contacts the amide nitrogen of Cys1 and the carbonyl oxygen of Pro-2 while His72, also of the TXXH motif, does not interact directly with the N-extein. Asp140 makes a water-mediated contact with the −1 carbonyl and a hydrogen bond with His147, also a conserved residue. Suc 159 is the C-terminal succinimide.
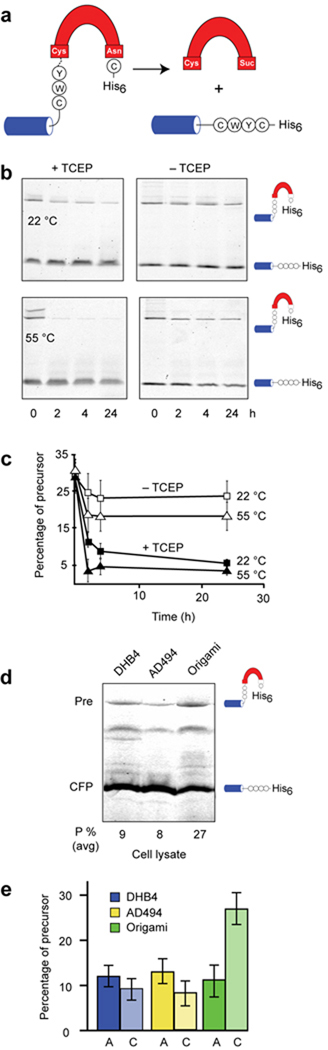
Protein splicing by the P. abssyi MoaA intein is redox sensitive (a). Schematic of CWYCMoaA precursor. Splicing of the MoaA intein (red) with native N-extein residues (circles) ligates the cyan fluorescent protein to a hexahistidine tag (His6). (b) and (c). Suppressed activity of the CWYCMoaA intein in the absence of a reducing agent. Intein autoprocessing was monitored in the presence and absence of the reducing agent, TCEP, at 22 °C (squares) and 55 °C (triangles). Precursor (CFPCWYCMoaA-His6) and splicing product (CFP-His6) were separated by nonreducing SDS-PAGE and detected by in-gel fluorescence (excitation 457 nm, emission 526 nm). Plot of precursor disappearance in panel c, using data of panel b, shows that TCEP is required for efficient CWYCMoaA processing at 22 °C and at 55 °C. Error bars represent s.d., n=3. (d). CWYCMoaA displays redox regulation in E. coli. Representative gel images showing CFP-fused MoaA precursor with wild-type Cys-3 (CFPCWYCMoaA-His6), and CFP-products remaining after 3 h of induction in DHB4 (left lane), AD494 (middle lane), and origami (right lane). (e). Cys-3 is required for MoaA precursor accumulation in origami. Graph was derived from images as in panel (a) and shows percent of unspliced CFPCWYCMoaA-His6 (C, light shaded) and CFPAWYCMoaA-His6 (A, dark shaded) precursors after expression in the indicated strains, averaged over three independent trials. Error bars represent s.d.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
