

Order of the proteasomal ATPases and eukaryotic proteasome assembly
- PMID: 21461838
- PMCID: PMC3256250
- DOI: 10.1007/s12013-011-9178-4
Order of the proteasomal ATPases and eukaryotic proteasome assembly
Abstract
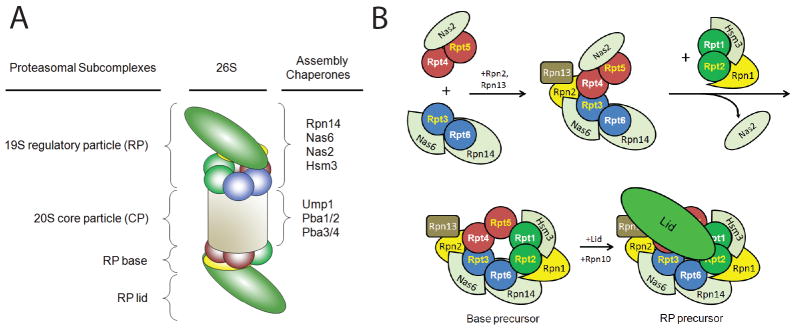
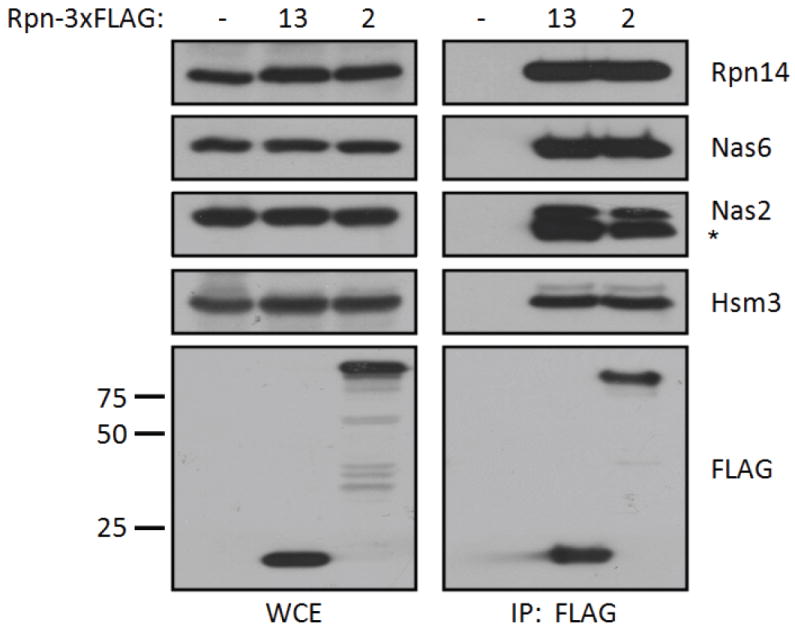
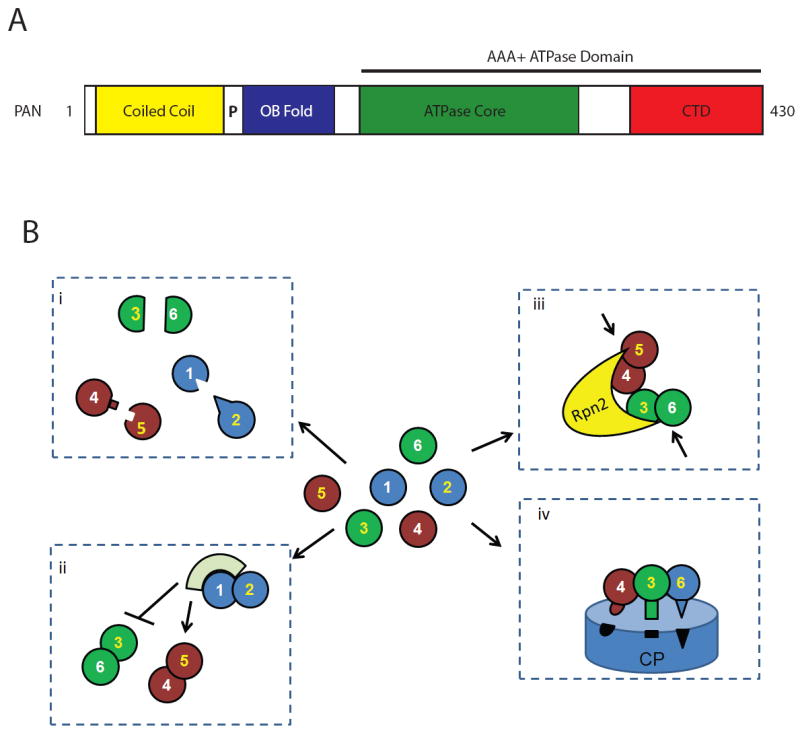
The 26S proteasome is responsible for a large fraction of the regulated protein degradation in eukaryotic cells. The enzyme complex is composed of a 20S proteolytic core particle (CP) capped on one or both ends with a 19S regulatory particle (RP). The RP recognizes and unfolds substrates and translocates them into the CP. The RP can be further divided into lid and base subcomplexes. The base contains a ring of six AAA+ ATPases (Rpts) that directly abuts the CP and is responsible for unfolding substrates and driving them into the CP for proteolysis. Although 120 arrangements of the six different ATPases within the ring are possible in principle, they array themselves in one specific order. The high sequence and structural similarity between the Rpt subunits presents special challenges for their ordered association and incorporation into the assembling proteasome. In this review, we discuss recent advances in our understanding of proteasomal RP base biogenesis, with emphasis on potential specificity determinants in ring arrangement, and the implications of the ATPase ring arrangement for proteasome assembly.
Figures
Similar articles
-
Heterohexameric ring arrangement of the eukaryotic proteasomal ATPases: implications for proteasome structure and assembly.Mol Cell. 2010 May 14;38(3):393-403. doi: 10.1016/j.molcel.2010.02.035. Mol Cell. 2010. PMID: 20471945 Free PMC article.
-
Proteasomal AAA-ATPases: structure and function.Biochim Biophys Acta. 2012 Jan;1823(1):67-82. doi: 10.1016/j.bbamcr.2011.07.009. Epub 2011 Jul 23. Biochim Biophys Acta. 2012. PMID: 21820014 Review.
-
Stable incorporation of ATPase subunits into 19 S regulatory particle of human proteasome requires nucleotide binding and C-terminal tails.J Biol Chem. 2012 Mar 16;287(12):9269-79. doi: 10.1074/jbc.M111.316208. Epub 2012 Jan 24. J Biol Chem. 2012. PMID: 22275368 Free PMC article.
-
Assembly manual for the proteasome regulatory particle: the first draft.Biochem Soc Trans. 2010 Feb;38(Pt 1):6-13. doi: 10.1042/BST0380006. Biochem Soc Trans. 2010. PMID: 20074027 Free PMC article. Review.
-
Multiple assembly chaperones govern biogenesis of the proteasome regulatory particle base.Cell. 2009 May 29;137(5):887-99. doi: 10.1016/j.cell.2009.04.061. Epub 2009 May 14. Cell. 2009. PMID: 19446322 Free PMC article.
Cited by
-
Incorporation of the Rpn12 subunit couples completion of proteasome regulatory particle lid assembly to lid-base joining.Mol Cell. 2011 Dec 23;44(6):907-17. doi: 10.1016/j.molcel.2011.11.020. Mol Cell. 2011. PMID: 22195964 Free PMC article.
-
Prokaryotic proteasomes: nanocompartments of degradation.J Mol Microbiol Biotechnol. 2013;23(4-5):321-34. doi: 10.1159/000351348. Epub 2013 Aug 5. J Mol Microbiol Biotechnol. 2013. PMID: 23920495 Free PMC article. Review.
-
Proteasome Structure and Assembly.J Mol Biol. 2017 Nov 10;429(22):3500-3524. doi: 10.1016/j.jmb.2017.05.027. Epub 2017 Jun 3. J Mol Biol. 2017. PMID: 28583440 Free PMC article. Review.
-
Structural basis for specific recognition of Rpt1p, an ATPase subunit of 26 S proteasome, by proteasome-dedicated chaperone Hsm3p.J Biol Chem. 2012 Apr 6;287(15):12172-82. doi: 10.1074/jbc.M112.345876. Epub 2012 Feb 8. J Biol Chem. 2012. PMID: 22334676 Free PMC article.
-
Fate of pup inside the Mycobacterium proteasome studied by in-cell NMR.PLoS One. 2013 Sep 10;8(9):e74576. doi: 10.1371/journal.pone.0074576. eCollection 2013. PLoS One. 2013. PMID: 24040288 Free PMC article.
References
-
- Kerscher O, Felberbaum R, Hochstrasser M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu Rev Cell Dev Biol. 2006;22:159–80. - PubMed
-
- Elsasser S, Finley D. Delivery of ubiquitinated substrates to protein-unfolding machines. Nat Cell Biol. 2005;7:742–9. - PubMed
-
- Pickart CM, Cohen RE. Proteasomes and their kin: proteases in the machine age. Nat Rev Mol Cell Biol. 2004;5:177–87. - PubMed
-
- Hochstrasser M, Wang J. Unraveling the means to the end in ATP-dependent proteases. Nat Struct Biol. 2001;8:294–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
