

Developmental plasticity of auditory cortical inhibitory synapses
- PMID: 21463668
- PMCID: PMC3135718
- DOI: 10.1016/j.heares.2011.03.015
Developmental plasticity of auditory cortical inhibitory synapses
Abstract
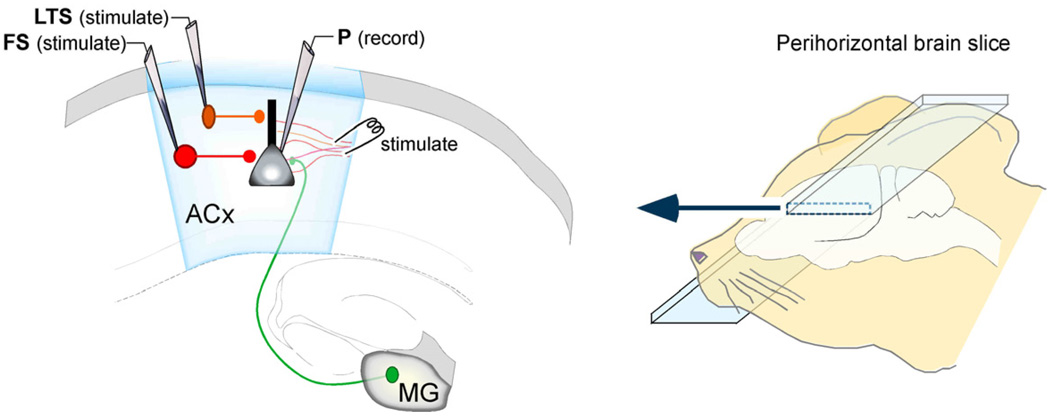
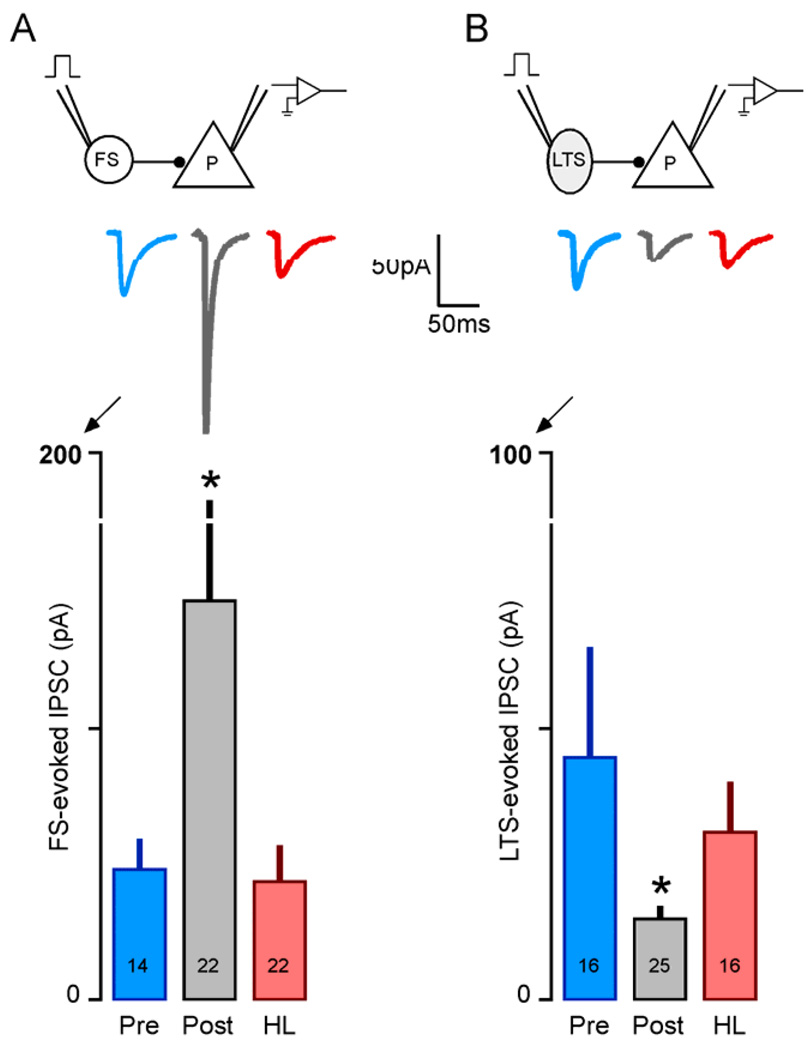
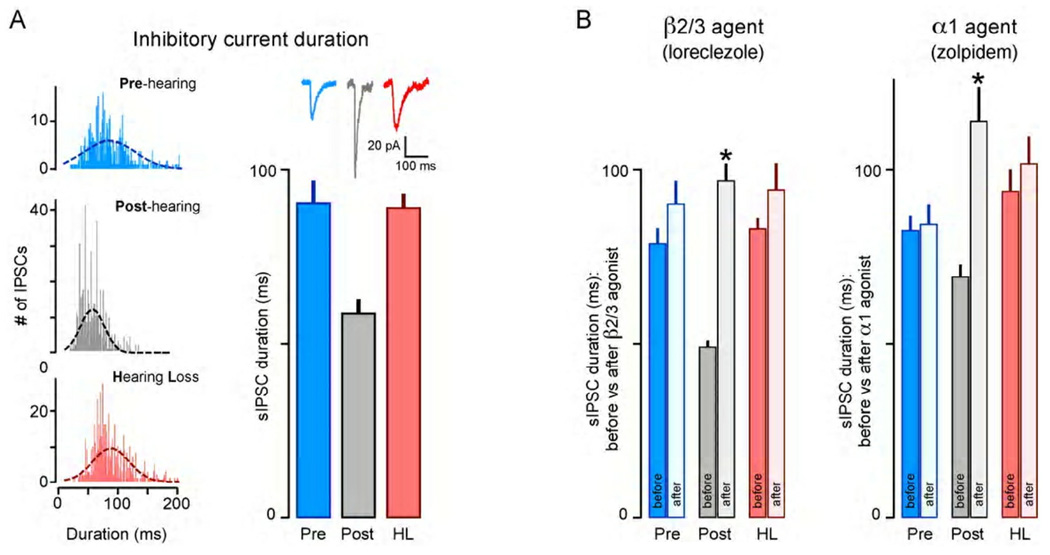
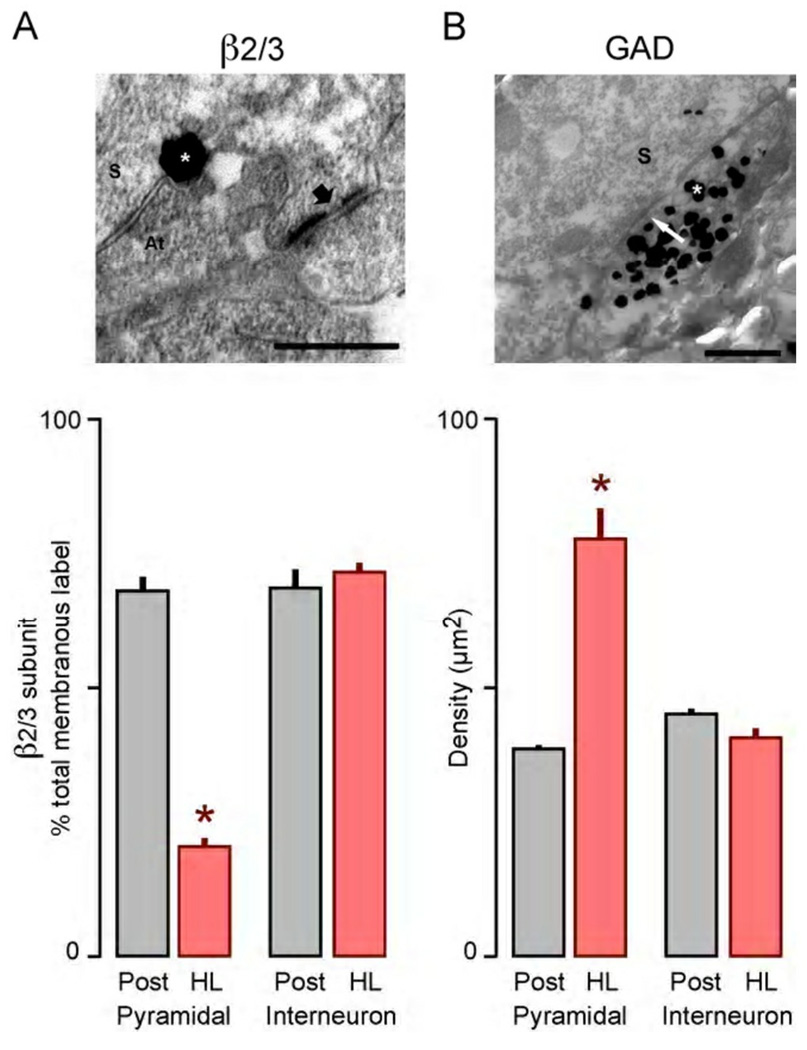
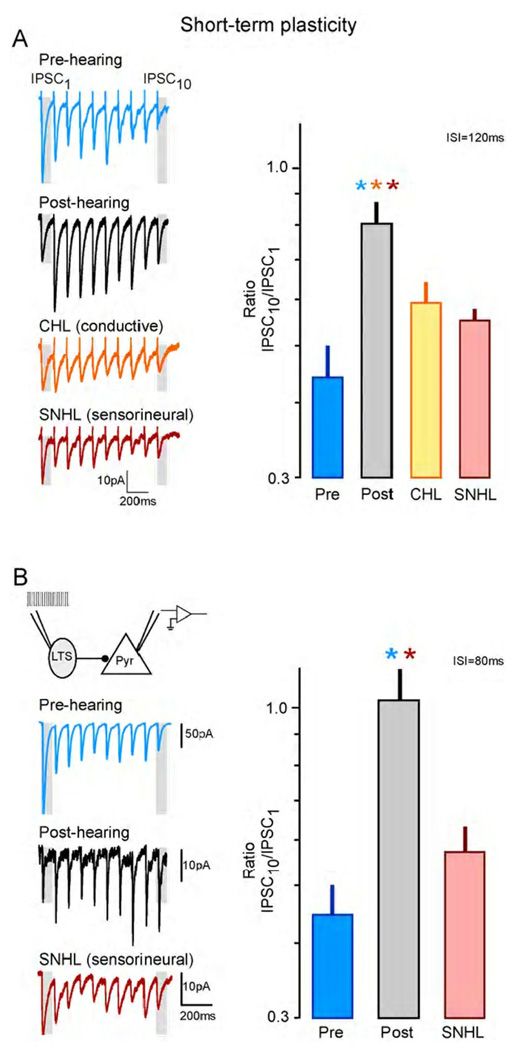
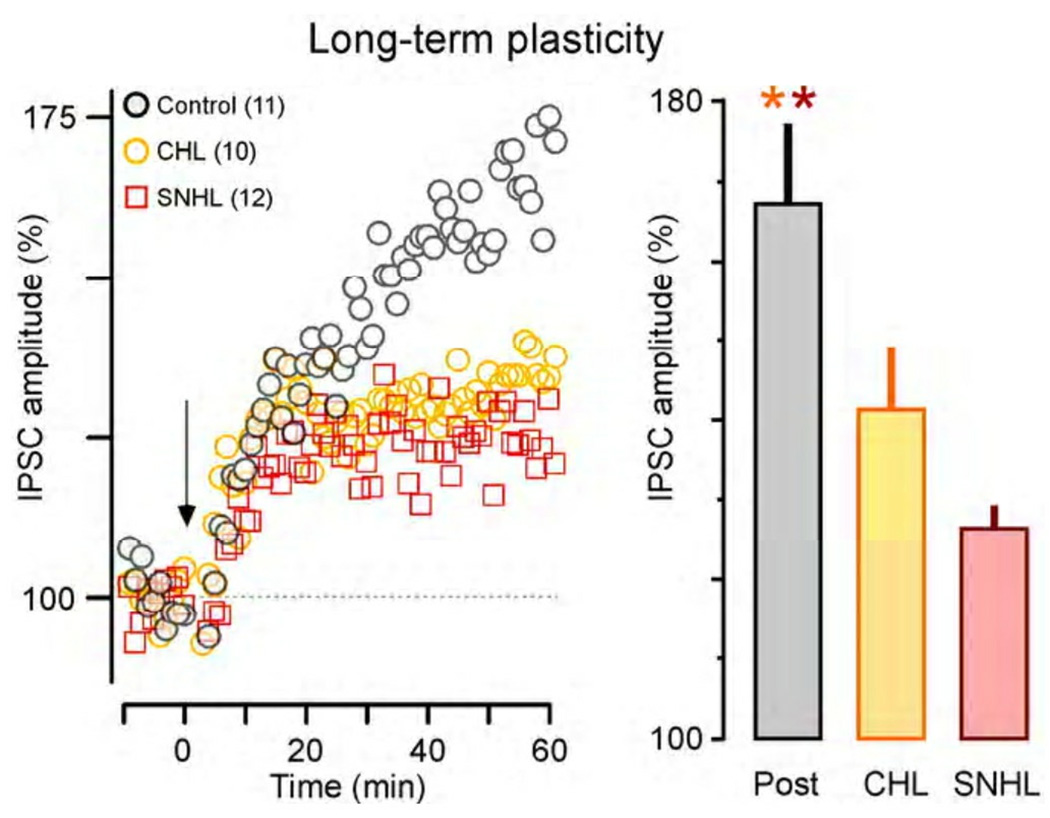
Functional inhibitory synapses form in auditory cortex well before the onset of normal hearing. However, their properties change dramatically during normal development, and many of these maturational events are delayed by hearing loss. Here, we review recent findings on the developmental plasticity of inhibitory synapse strength, kinetics, and GABAA receptor localization in auditory cortex. Although hearing loss generally leads to a reduction of inhibitory strength, this depends on the type of presynaptic interneuron. Furthermore, plasticity of inhibitory synapses also depends on the postsynaptic target. Hearing loss leads reduced GABAA receptor localization to the membrane of excitatory, but not inhibitory neurons. A reduction in normal activity in development can also affect the use-dependent plasticity of inhibitory synapses. Even moderate hearing loss can disrupt inhibitory short- and long-term synaptic plasticity. Thus, the cortex did not compensate for the loss of inhibition in the brainstem, but rather exacerbated the response to hearing loss by further reducing inhibitory drive. Together, these results demonstrate that inhibitory synapses are exceptionally dynamic during development, and deafness-induced perturbation of inhibitory properties may have a profound impact on auditory processing.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
References
-
- Abraham WC, Bear MF. Metaplasticity: the plasticity of synaptic plasticity. Trends Neurosci. 1996;19:126–130. - PubMed
-
- Bacon SP, Gleitman RM. Modulation detection in subjects with relatively flat hearing losses. J Speech Hear Res. 1992;35:642–653. - PubMed
-
- Banks MI, Hardie JB, Pearce RA. Development of GABA(A) receptor-mediated inhibitory postsynaptic currents in hippocampus. J Neurophysiol. 2002;88:3097–3107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
