

The Agr-like quorum-sensing system regulates sporulation and production of enterotoxin and beta2 toxin by Clostridium perfringens type A non-food-borne human gastrointestinal disease strain F5603
- PMID: 21464088
- PMCID: PMC3125842
- DOI: 10.1128/IAI.00169-11
The Agr-like quorum-sensing system regulates sporulation and production of enterotoxin and beta2 toxin by Clostridium perfringens type A non-food-borne human gastrointestinal disease strain F5603
Abstract
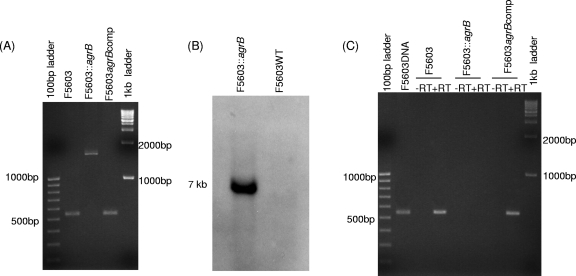
Clostridium perfringens type A strains producing enterotoxin (CPE) cause one of the most common bacterial food-borne illnesses, as well as many cases of non-food-borne human gastrointestinal disease. Recent studies have shown that an Agr-like quorum-sensing system controls production of chromosomally encoded alpha-toxin and perfringolysin O by C. perfringens, as well as sporulation by Clostridium botulinum and Clostridium sporogenes. The current study explored whether the Agr-like quorum-sensing system also regulates sporulation and production of two plasmid-encoded toxins (CPE and beta2 toxin) that may contribute to the pathogenesis of non-food-borne human gastrointestinal disease strain F5603. An isogenic agrB null mutant was inhibited for production of beta2 toxin during vegetative growth and in sporulating culture, providing the first evidence that, in C. perfringens, this system can control production of plasmid-encoded toxins as well as chromosomally encoded toxins. This mutant also showed reduced production of alpha-toxin and perfringolysin O during vegetative growth. Importantly, when cultured in sporulation medium, the mutant failed to efficiently form spores and was blocked for CPE production. Complementation partially or fully reversed all phenotypic changes in the mutant, confirming that they were specifically due to inactivation of the agr locus. Western blots suggest that this loss of sporulation and sporulation-specific CPE production for the agrB null mutant involves, at least in part, Agr-mediated regulation of production of Spo0A and alternative sigma factors, which are essential for C. perfringens sporulation.
Figures

Similar articles
-
Identification of an Important Orphan Histidine Kinase for the Initiation of Sporulation and Enterotoxin Production by Clostridium perfringens Type F Strain SM101.mBio. 2019 Jan 22;10(1):e02674-18. doi: 10.1128/mBio.02674-18. mBio. 2019. PMID: 30670619 Free PMC article.
-
NanR Regulates Sporulation and Enterotoxin Production by Clostridium perfringens Type F Strain F4969.Infect Immun. 2018 Sep 21;86(10):e00416-18. doi: 10.1128/IAI.00416-18. Print 2018 Oct. Infect Immun. 2018. PMID: 30082481 Free PMC article.
-
CodY Promotes Sporulation and Enterotoxin Production by Clostridium perfringens Type A Strain SM101.Infect Immun. 2017 Feb 23;85(3):e00855-16. doi: 10.1128/IAI.00855-16. Print 2017 Mar. Infect Immun. 2017. PMID: 28052992 Free PMC article.
-
The biology and pathogenicity of Clostridium perfringens type F: a common human enteropathogen with a new(ish) name.Microbiol Mol Biol Rev. 2024 Sep 26;88(3):e0014023. doi: 10.1128/mmbr.00140-23. Epub 2024 Jun 12. Microbiol Mol Biol Rev. 2024. PMID: 38864615 Free PMC article. Review.
-
Host cell-induced signaling causes Clostridium perfringens to upregulate production of toxins important for intestinal infections.Gut Microbes. 2014 Jan-Feb;5(1):96-107. doi: 10.4161/gmic.26419. Epub 2013 Sep 10. Gut Microbes. 2014. PMID: 24061146 Free PMC article. Review.
Cited by
-
Quorum Sensing Inhibitors: An Alternative Strategy to Win the Battle against Multidrug-Resistant (MDR) Bacteria.Molecules. 2024 Jul 24;29(15):3466. doi: 10.3390/molecules29153466. Molecules. 2024. PMID: 39124871 Free PMC article. Review.
-
Targeting agr- and agr-Like quorum sensing systems for development of common therapeutics to treat multiple gram-positive bacterial infections.Sensors (Basel). 2013 Apr 18;13(4):5130-66. doi: 10.3390/s130405130. Sensors (Basel). 2013. PMID: 23598501 Free PMC article. Review.
-
Characterization of thermostable bacteriophage CPD2 and its endolysin LysCPD2 as biocontrol agents against Clostridium perfringens.Food Sci Biotechnol. 2023 May 24;32(14):2069-2077. doi: 10.1007/s10068-023-01314-0. eCollection 2023 Dec. Food Sci Biotechnol. 2023. PMID: 37860732 Free PMC article.
-
Identification of orphan histidine kinases that impact sporulation and enterotoxin production by Clostridium perfringens type F strain SM101 in a pathophysiologically-relevant ex vivo mouse intestinal contents model.PLoS Pathog. 2023 Jun 1;19(6):e1011429. doi: 10.1371/journal.ppat.1011429. eCollection 2023 Jun. PLoS Pathog. 2023. PMID: 37262083 Free PMC article.
-
Enterotoxic Clostridia: Clostridium perfringens Enteric Diseases.Microbiol Spectr. 2018 Sep;6(5):10.1128/microbiolspec.gpp3-0003-2017. doi: 10.1128/microbiolspec.GPP3-0003-2017. Microbiol Spectr. 2018. PMID: 30238869 Free PMC article.
References
-
- Carman R. J. 1997. Clostridium perfringens in spontaneous and antibiotic-associated diarrhoea of man and other animals. Rev. Med. Microbiol. 8(Suppl. 1):S43–S45
-
- Collie R. E., Kokai-Kun J. F., McClane B. A. 1998. Phenotypic characterization of enterotoxigenic Clostridium perfringens isolates from non-food-borne human gastrointestinal diseases. Anaerobe 4:69–79 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
