

Gdnf is mitogenic, neurotrophic, and chemoattractive to enteric neural crest cells in the embryonic colon
- PMID: 21465624
- PMCID: PMC3092856
- DOI: 10.1002/dvdy.22630
Gdnf is mitogenic, neurotrophic, and chemoattractive to enteric neural crest cells in the embryonic colon
Abstract
Glial-derived neurotrophic factor (Gdnf) is required for morphogenesis of the enteric nervous system (ENS) and it has been shown to regulate proliferation, differentiation, and survival of cultured enteric neural crest-derived cells (ENCCs). The goal of this study was to investigate its in vivo role in the colon, the site most commonly affected by intestinal neuropathies such as Hirschsprung's disease. Gdnf activity was modulated in ovo in the distal gut of avian embryos using targeted retrovirus-mediated gene overexpression and retroviral vector-based gene silencing. We find that Gdnf has a pleiotropic effect on colonic ENCCs, promoting proliferation, inducing neuronal differentiation, and acting as a chemoattractant. Down-regulating Gdnf similarly induces premature neuronal differentiation, but also inhibits ENCC proliferation, leading to distal colorectal aganglionosis with severe proximal hypoganglionosis. These results indicate an important role for Gdnf signaling in colonic ENS formation and emphasize the critical balance between proliferation and differentiation in the developing ENS.
Copyright © 2011 Wiley-Liss, Inc.
Figures

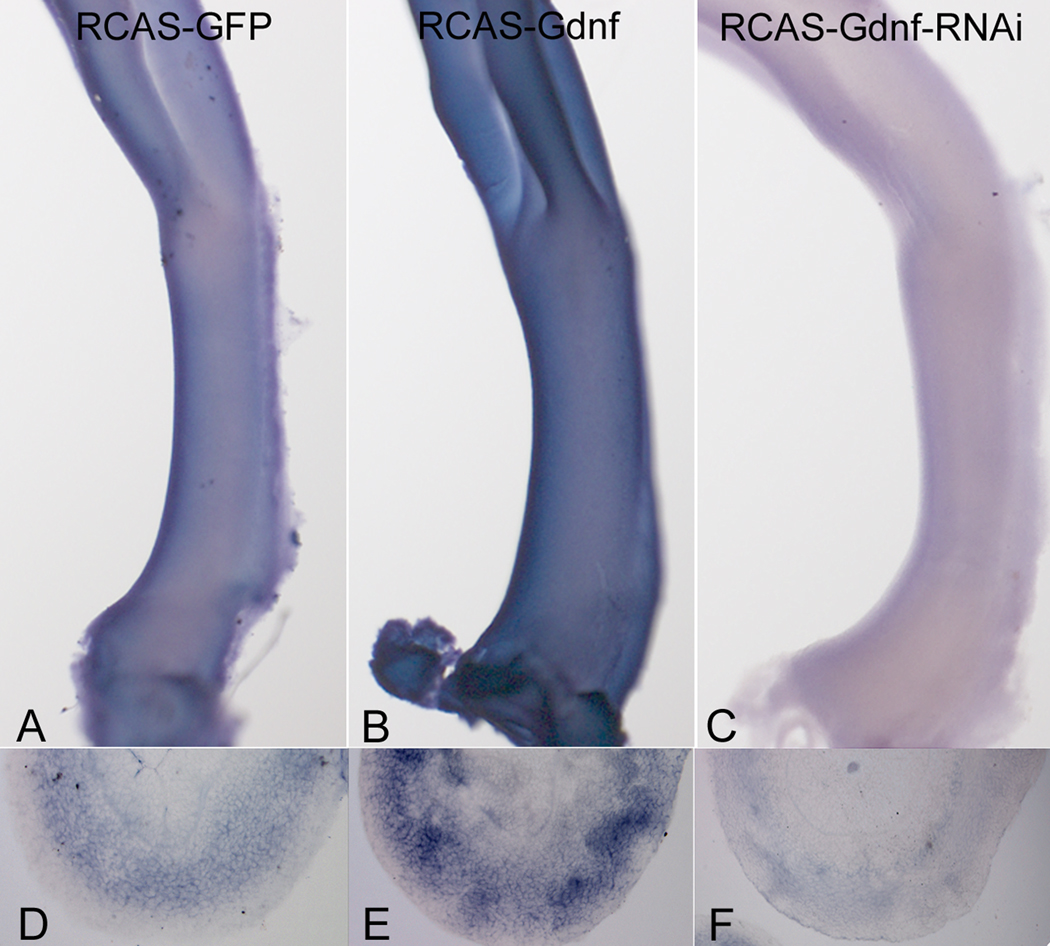

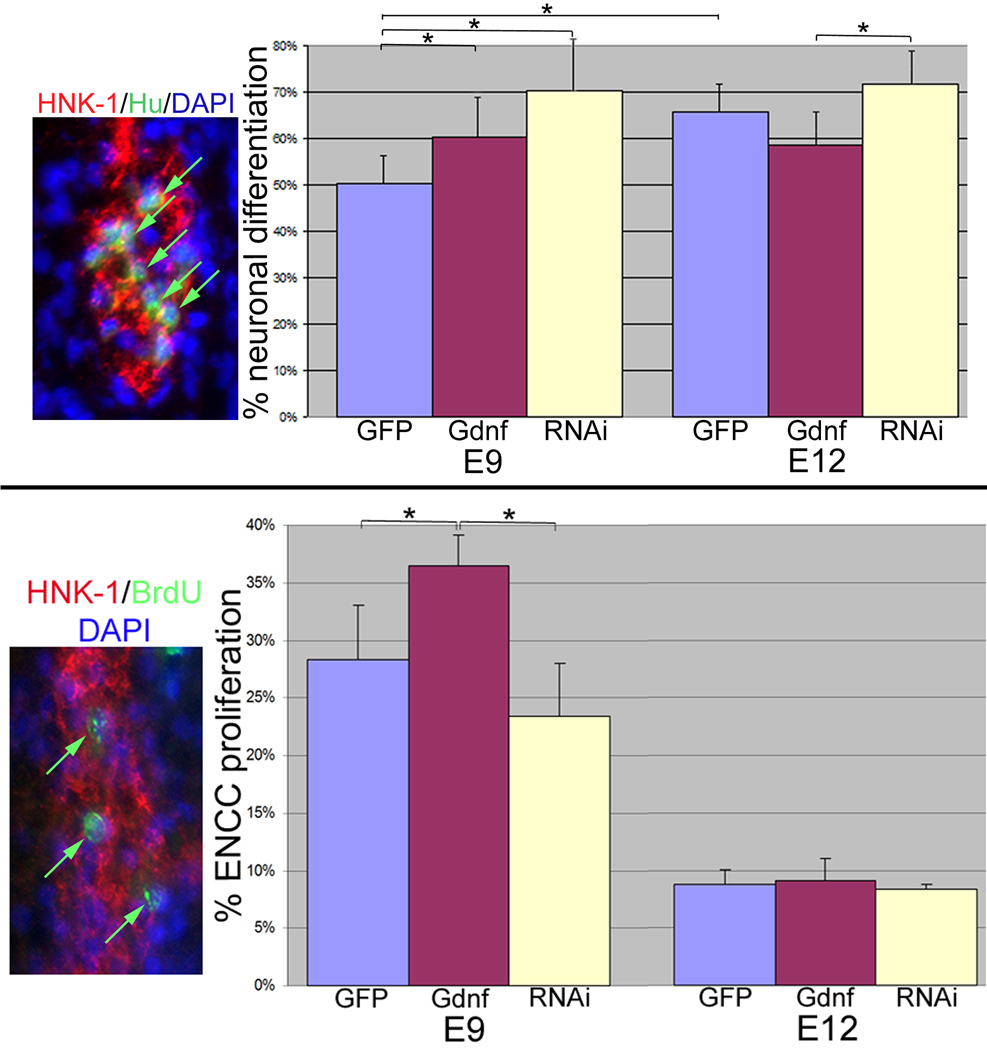
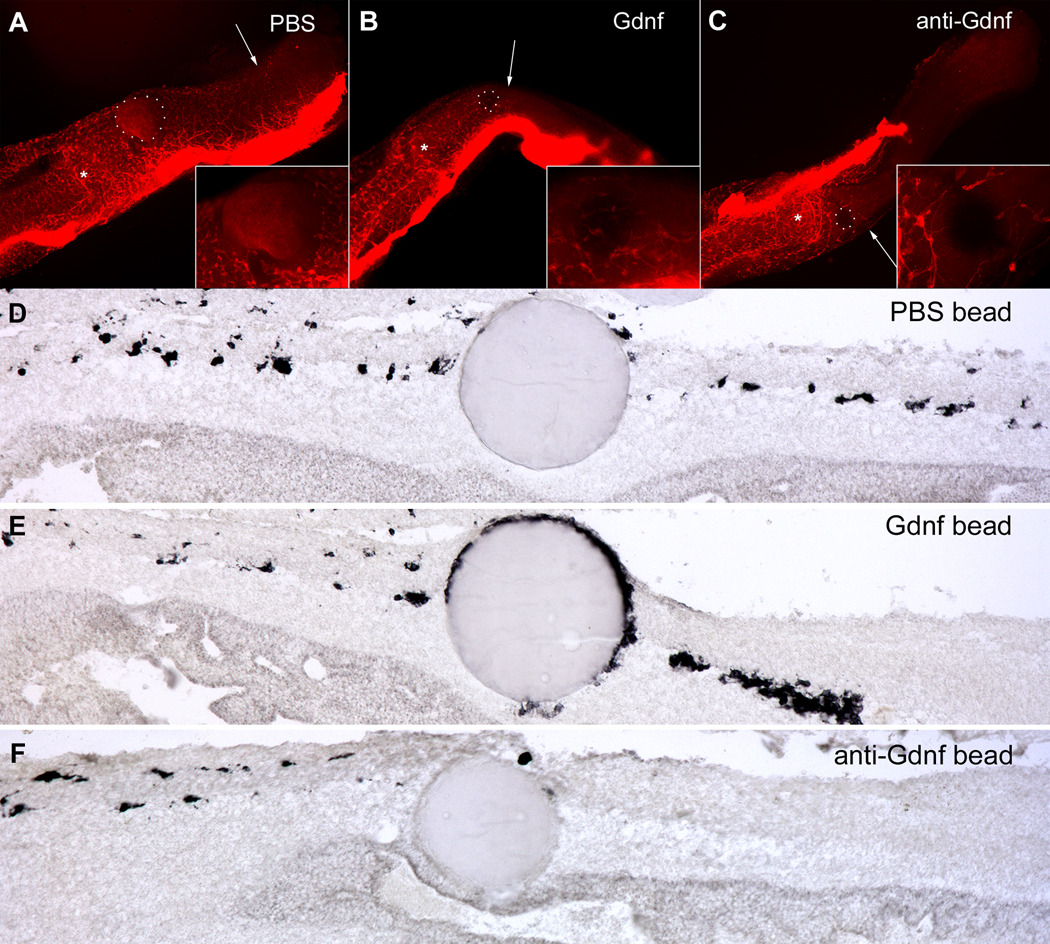
References
-
- Barlow A, de Graaff E, Pachnis V. Enteric nervous system progenitors are coordinately controlled by the G protein-coupled receptor EDNRB and the receptor tyrosine kinase RET. Neuron. 2003;40:905–916. - PubMed
-
- Barlow AJ, Wallace AS, Thapar N, Burns AJ. Critical numbers of neural crest cells are required in the pathways from the neural tube to the foregut to ensure complete enteric nervous system formation. Development. 2008;135:1681–1691. - PubMed
-
- Burns AJ, Le Douarin NM. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 1998;125:4335–4347. - PubMed
-
- Cepko CL. Transduction of genes using retroviral vectors. In: Ausubel RB FM, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K, editors. Current Protocols in Molecular Biology. New York: John Wiley and Sons; 1991. pp. 9:9.10.11–19.14.13.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
