

Molecular structures and interactions in the yeast kinetochore
- PMID: 21467141
- PMCID: PMC3207249
- DOI: 10.1101/sqb.2010.75.040
Molecular structures and interactions in the yeast kinetochore
Abstract
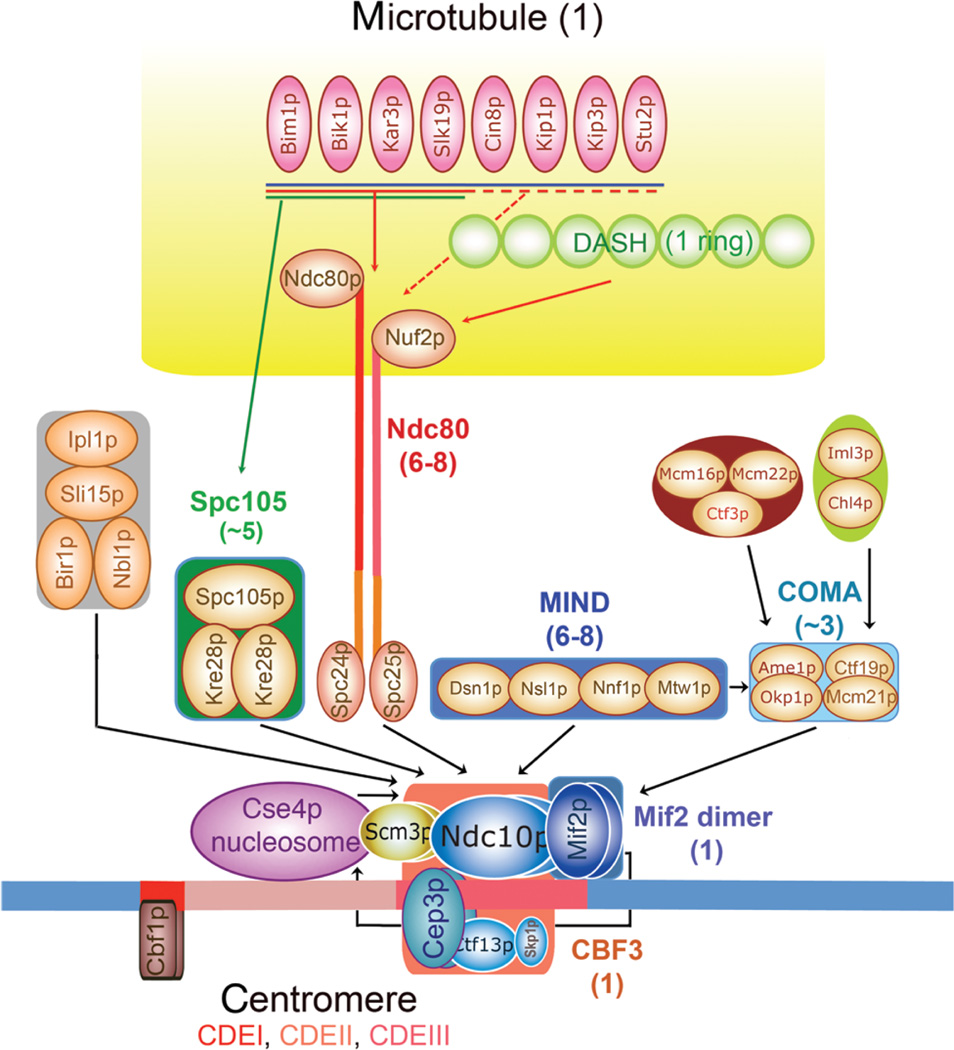
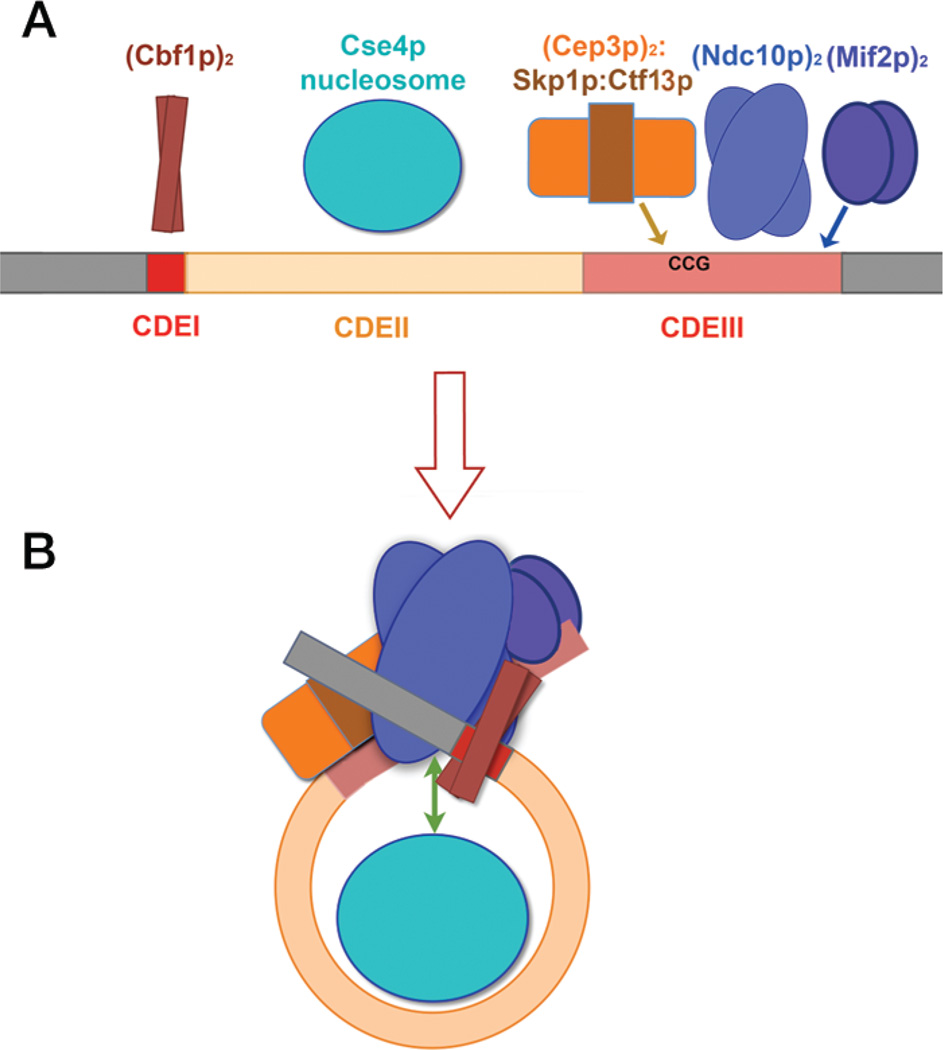
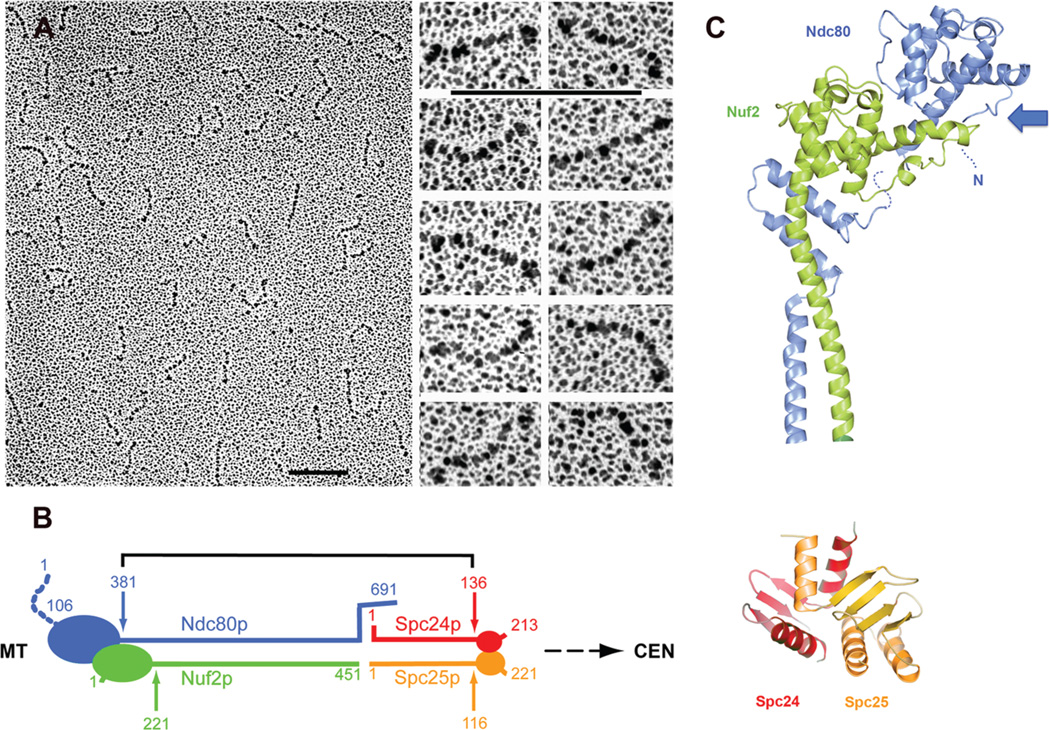
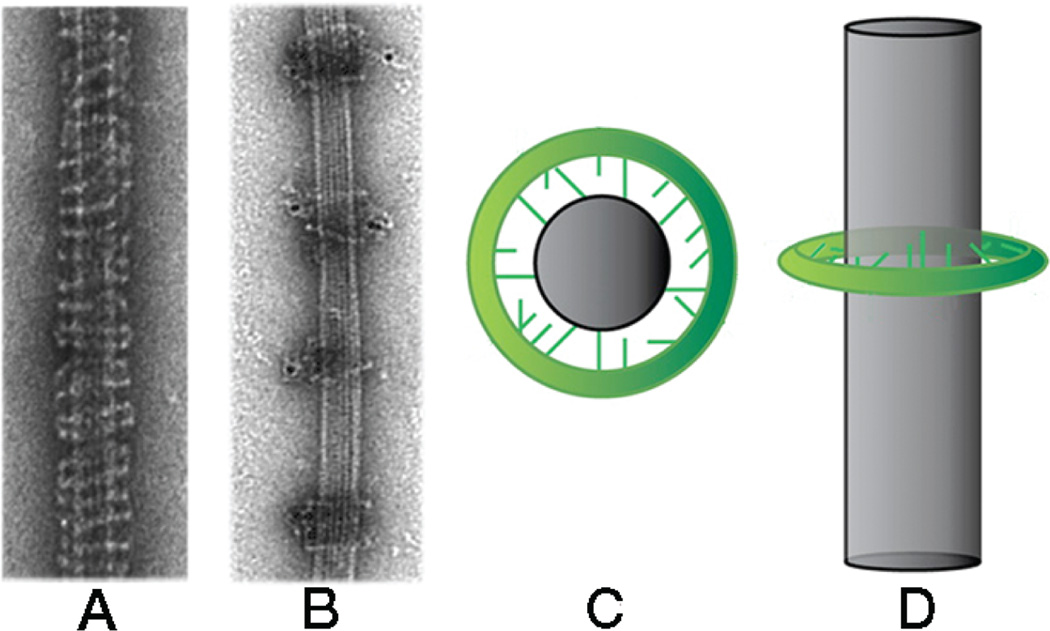
Kinetochores are the elaborate protein assemblies that attach chromosomes to spindle microtubules in mitosis and meiosis. The kinetochores of point-centromere yeast appear to represent an elementary module, which repeats a number of times in kinetochores assembled on regional centromeres. Structural analyses of the discrete protein subcomplexes that make up the budding-yeast kinetochore have begun to reveal principles of kinetochore architecture and to uncover molecular mechanisms underlying functions such as transmission of tension and establishment and maintenance of bipolar attachment. The centromeric DNA is probably wrapped into a compact organization, not only by a conserved, centromeric nucleosome, but also by interactions among various other DNA-bound kinetochore components. The rod-like, heterotetrameric Ndc80 complex, roughly 600 Å long, appears to extend from the DNA-proximal assembly to the plus end of a microtubule, to which one end of the complex is known to bind. Ongoing structural studies will clarify the roles of a number of other well-defined complexes.
Figures

Similar articles
-
Structure of the inner kinetochore CCAN complex assembled onto a centromeric nucleosome.Nature. 2019 Oct;574(7777):278-282. doi: 10.1038/s41586-019-1609-1. Epub 2019 Oct 2. Nature. 2019. PMID: 31578520 Free PMC article.
-
Molecular architecture and connectivity of the budding yeast Mtw1 kinetochore complex.J Mol Biol. 2011 Jan 14;405(2):548-59. doi: 10.1016/j.jmb.2010.11.012. Epub 2010 Nov 12. J Mol Biol. 2011. PMID: 21075115 Free PMC article.
-
Molecular organization of the Ndc80 complex, an essential kinetochore component.Proc Natl Acad Sci U S A. 2005 Apr 12;102(15):5363-7. doi: 10.1073/pnas.0501168102. Epub 2005 Apr 4. Proc Natl Acad Sci U S A. 2005. PMID: 15809444 Free PMC article.
-
Structures and functions of yeast kinetochore complexes.Annu Rev Biochem. 2007;76:563-91. doi: 10.1146/annurev.biochem.76.052705.160607. Annu Rev Biochem. 2007. PMID: 17362199 Review.
-
The Ndc80 complex: hub of kinetochore activity.FEBS Lett. 2007 Jun 19;581(15):2862-9. doi: 10.1016/j.febslet.2007.05.012. Epub 2007 May 11. FEBS Lett. 2007. PMID: 17521635 Review.
Cited by
-
Structure of the MIND Complex Defines a Regulatory Focus for Yeast Kinetochore Assembly.Cell. 2016 Nov 3;167(4):1014-1027.e12. doi: 10.1016/j.cell.2016.10.011. Epub 2016 Oct 27. Cell. 2016. PMID: 27881300 Free PMC article.
-
Pat1 protects centromere-specific histone H3 variant Cse4 from Psh1-mediated ubiquitination.Mol Biol Cell. 2015 Jun 1;26(11):2067-79. doi: 10.1091/mbc.E14-08-1335. Epub 2015 Apr 1. Mol Biol Cell. 2015. PMID: 25833709 Free PMC article.
-
Budding yeast kinetochore proteins, Chl4 and Ctf19, are required to maintain SPB-centromere proximity during G1 and late anaphase.PLoS One. 2014 Jul 8;9(7):e101294. doi: 10.1371/journal.pone.0101294. eCollection 2014. PLoS One. 2014. PMID: 25003500 Free PMC article.
-
Structure, assembly and reading of centromeric chromatin.Curr Opin Genet Dev. 2012 Apr;22(2):139-47. doi: 10.1016/j.gde.2011.11.005. Epub 2011 Dec 16. Curr Opin Genet Dev. 2012. PMID: 22178421 Free PMC article. Review.
-
Structural integrity of centromeric chromatin and faithful chromosome segregation requires Pat1.Genetics. 2013 Oct;195(2):369-79. doi: 10.1534/genetics.113.155291. Epub 2013 Jul 26. Genetics. 2013. PMID: 23893485 Free PMC article.
References
-
- Camahort R, Li B, Florens L, Swanson SK, Washburn MP, Gerton JL. Scm3 is essential to recruit the histone h3 variant cse4 to centromeres and to maintain a functional kinetochore. Mol Cell. 2007;26:853–865. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases