

Poly(ADP-ribose) (PAR) binding to apoptosis-inducing factor is critical for PAR polymerase-1-dependent cell death (parthanatos)
- PMID: 21467298
- PMCID: PMC3086524
- DOI: 10.1126/scisignal.2000902
Poly(ADP-ribose) (PAR) binding to apoptosis-inducing factor is critical for PAR polymerase-1-dependent cell death (parthanatos)
Abstract
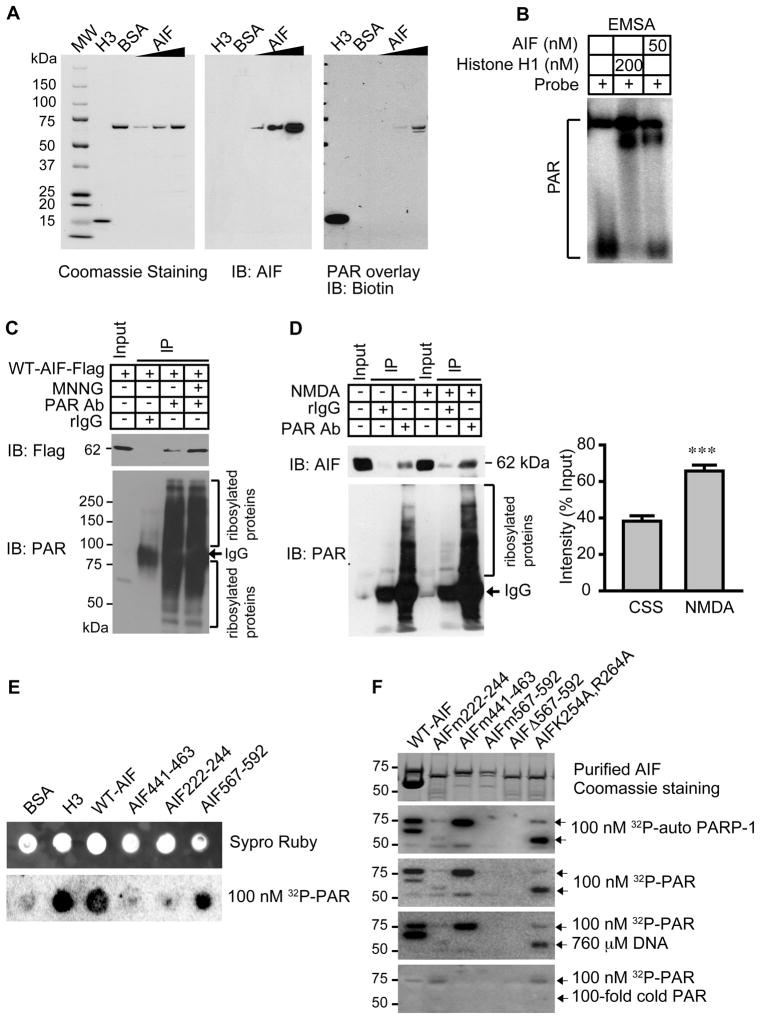
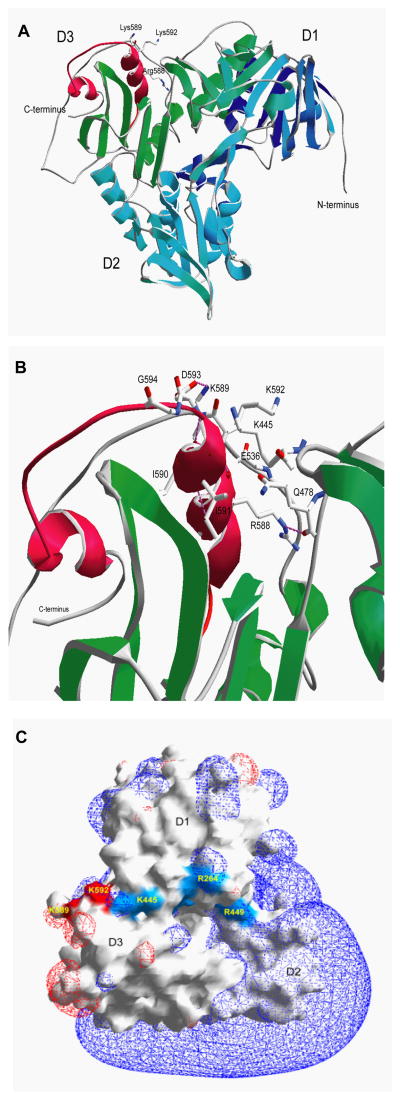
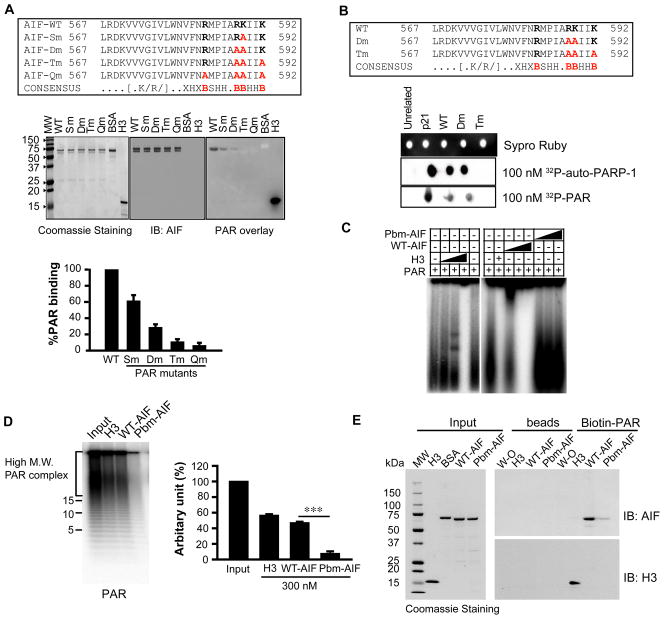
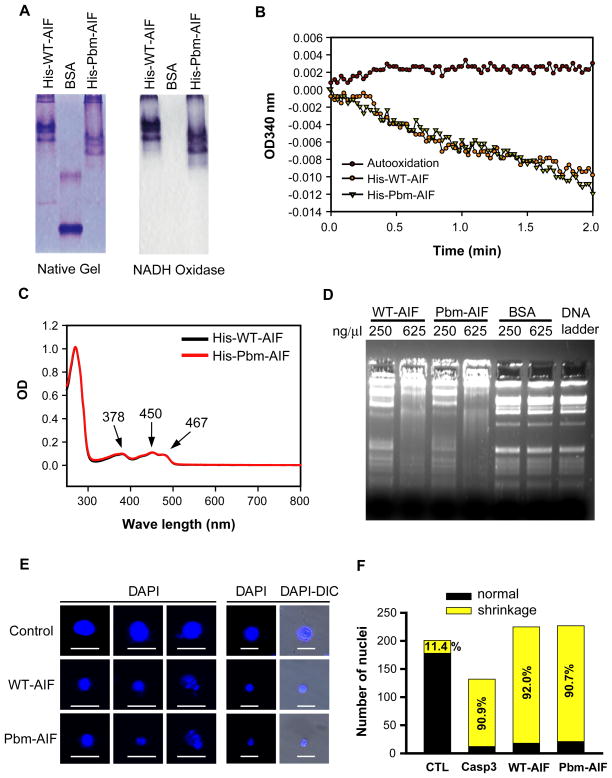
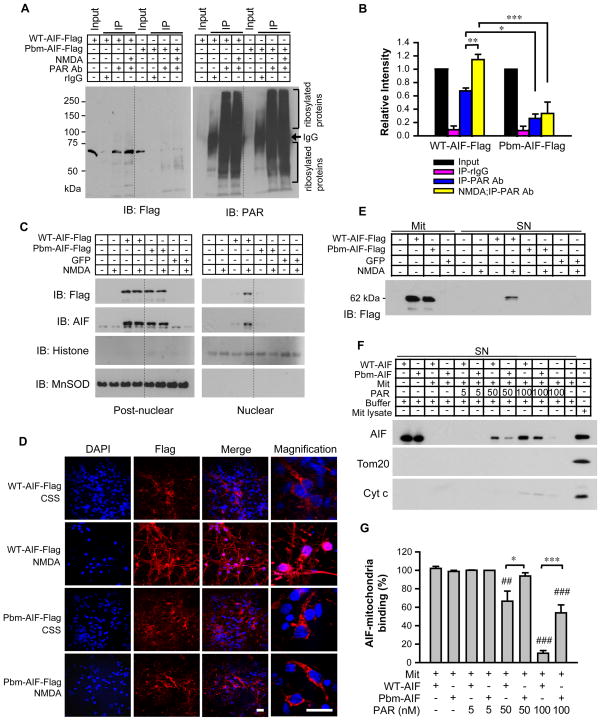
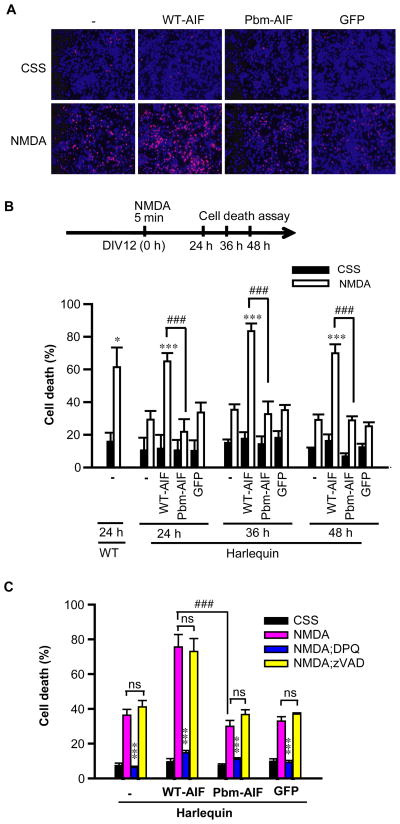
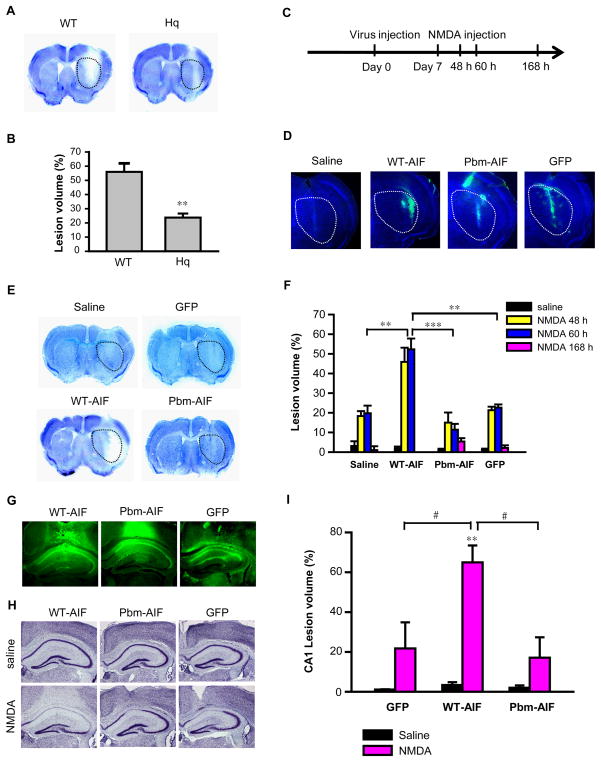
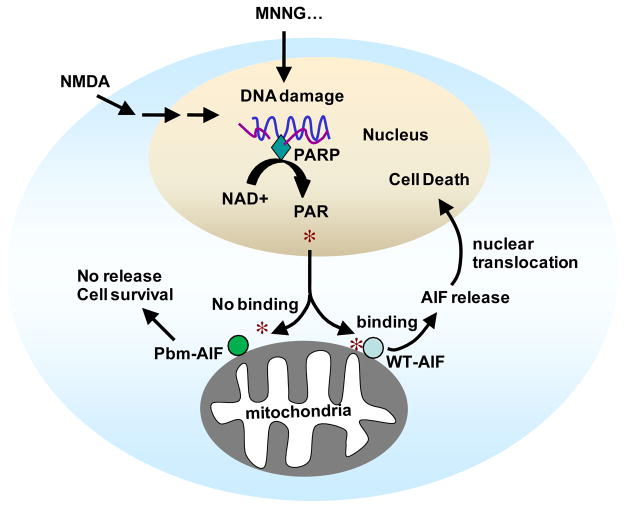
The mitochondrial protein apoptosis-inducing factor (AIF) plays a pivotal role in poly(ADP-ribose) polymerase-1 (PARP-1)-mediated cell death (parthanatos), during which it is released from the mitochondria and translocates to the nucleus. We show that AIF is a high-affinity poly(ADP-ribose) (PAR)-binding protein and that PAR binding to AIF is required for parthanatos both in vitro and in vivo. AIF bound PAR at a site distinct from AIF's DNA binding site, and this interaction triggered AIF release from the cytosolic side of the mitochondrial outer membrane. Mutation of the PAR binding site in AIF did not affect its NADH (reduced form of nicotinamide adenine dinucleotide) oxidase activity, its ability to bind FAD (flavin adenine dinucleotide) or DNA, or its ability to induce nuclear condensation. However, this AIF mutant was not released from mitochondria and did not translocate to the nucleus or mediate cell death after PARP-1 activation. These results suggest a mechanism for PARP-1 to initiate AIF-mediated cell death and indicate that AIF's bioenergetic cell survival-promoting functions are separate from its effects as a mitochondrially derived death effector. Interference with the PAR-AIF interaction or PAR signaling may provide notable opportunities for preventing cell death after activation of PARP-1.
Conflict of interest statement
Figures
References
-
- Cheung EC, Joza N, Steenaart NA, McClellan KA, Neuspiel M, McNamara S, MacLaurin JG, Rippstein P, Park DS, Shore GC, McBride HM, Penninger JM, Slack RS. Dissociating the dual roles of apoptosis-inducing factor in maintaining mitochondrial structure and apoptosis. Embo J. 2006;25:4061–4073. - PMC - PubMed
-
- Kang YH, Yi MJ, Kim MJ, Park MT, Bae S, Kang CM, Cho CK, Park IC, Park MJ, Rhee CH, Hong SI, Chung HY, Lee YS, Lee SJ. Caspase-independent cell death by arsenic trioxide in human cervical cancer cells: reactive oxygen species-mediated poly(ADP-ribose) polymerase-1 activation signals apoptosis-inducing factor release from mitochondria. Cancer Res. 2004;64:8960–8967. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
