

A structural and mutagenic blueprint for molecular recognition of strychnine and d-tubocurarine by different cys-loop receptors
- PMID: 21468359
- PMCID: PMC3066128
- DOI: 10.1371/journal.pbio.1001034
A structural and mutagenic blueprint for molecular recognition of strychnine and d-tubocurarine by different cys-loop receptors
Abstract
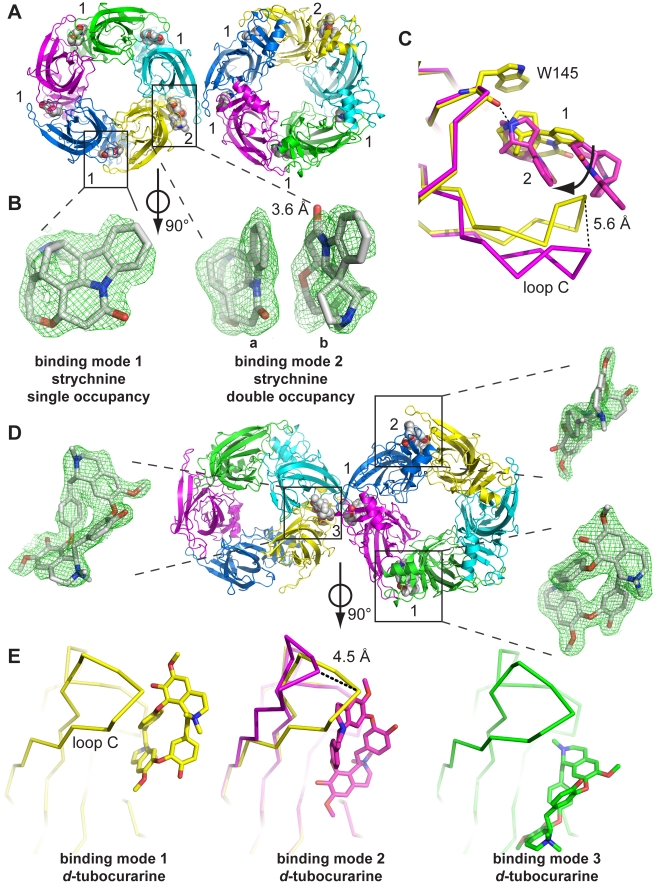
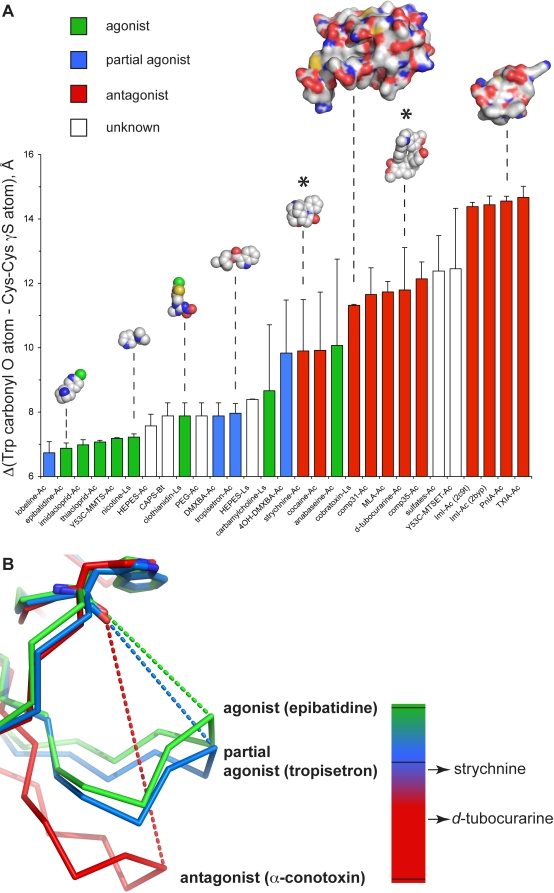
Cys-loop receptors (CLR) are pentameric ligand-gated ion channels that mediate fast excitatory or inhibitory transmission in the nervous system. Strychnine and d-tubocurarine (d-TC) are neurotoxins that have been highly instrumental in decades of research on glycine receptors (GlyR) and nicotinic acetylcholine receptors (nAChR), respectively. In this study we addressed the question how the molecular recognition of strychnine and d-TC occurs with high affinity and yet low specificity towards diverse CLR family members. X-ray crystal structures of the complexes with AChBP, a well-described structural homolog of the extracellular domain of the nAChRs, revealed that strychnine and d-TC adopt multiple occupancies and different ligand orientations, stabilizing the homopentameric protein in an asymmetric state. This introduces a new level of structural diversity in CLRs. Unlike protein and peptide neurotoxins, strychnine and d-TC form a limited number of contacts in the binding pocket of AChBP, offering an explanation for their low selectivity. Based on the ligand interactions observed in strychnine- and d-TC-AChBP complexes we performed alanine-scanning mutagenesis in the binding pocket of the human α1 GlyR and α7 nAChR and showed the functional relevance of these residues in conferring high potency of strychnine and d-TC, respectively. Our results demonstrate that a limited number of ligand interactions in the binding pocket together with an energetic stabilization of the extracellular domain are key to the poor selective recognition of strychnine and d-TC by CLRs as diverse as the GlyR, nAChR, and 5-HT(3)R.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Papineni R. V, Pedersen S. E. Interaction of d-tubocurarine analogs with the mouse nicotinic acetylcholine receptor. Ligand orientation at the binding site. J Biol Chem. 1997;272:24891–24898. - PubMed
-
- Jonsson M, Gurley D, Dabrowski M, Larsson O, Johnson E. C, et al. Distinct pharmacologic properties of neuromuscular blocking agents on human neuronal nicotinic acetylcholine receptors: a possible explanation for the train-of-four fade. Anesthesiology. 2006;105:521–533. - PubMed
-
- Hope A. G, Belelli D, Mair I. D, Lambert J. J, Peters J. A. Molecular determinants of (+)-tubocurarine binding at recombinant 5-hydroxytryptamine3A receptor subunits. Mol Pharmacol. 1999;55:1037–1043. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
