

Rescue of infectious particles from preassembled alphavirus nucleocapsid cores
- PMID: 21471237
- PMCID: PMC3126313
- DOI: 10.1128/JVI.00039-11
Rescue of infectious particles from preassembled alphavirus nucleocapsid cores
Abstract
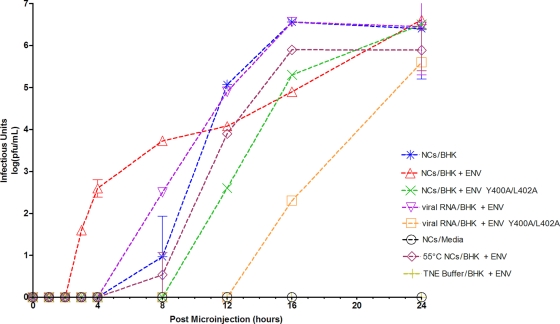
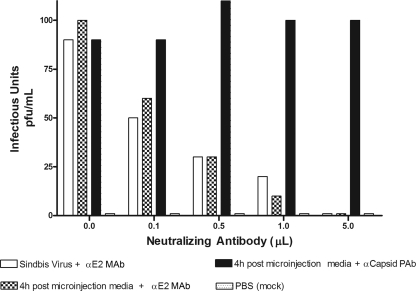
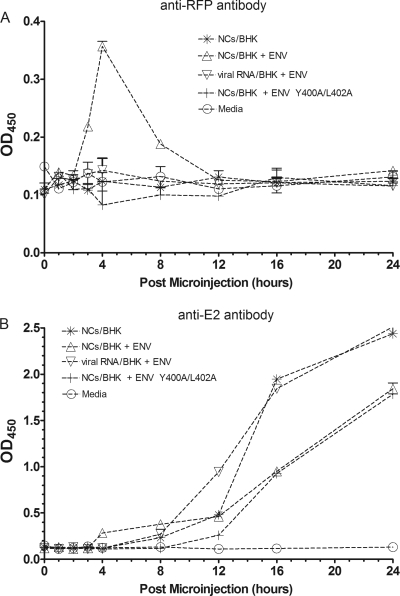
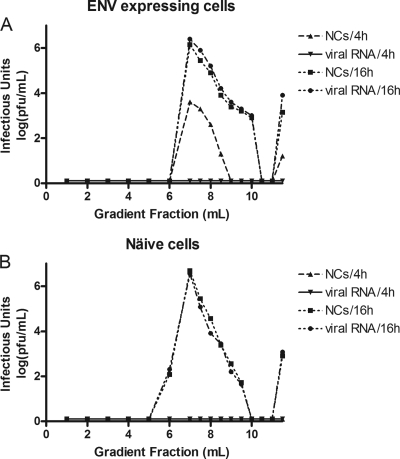
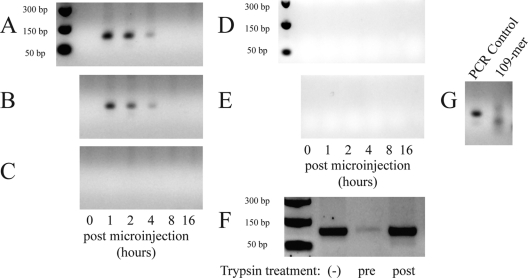
Alphaviruses are small, spherical, enveloped, positive-sense, single-stranded, RNA viruses responsible for considerable human and animal disease. Using microinjection of preassembled cores as a tool, a system has been established to study the assembly and budding process of Sindbis virus, the type member of the alphaviruses. We demonstrate the release of infectious virus-like particles from cells expressing Sindbis virus envelope glycoproteins following microinjection of Sindbis virus nucleocapsids purified from the cytoplasm of infected cells. Furthermore, it is shown that nucleocapsids assembled in vitro mimic those isolated in the cytoplasm of infected cells with respect to their ability to be incorporated into enveloped virions following microinjection. This system allows for the study of the alphavirus budding process independent of an authentic infection and provides a platform to study viral and host requirements for budding.
Figures
References
-
- Garoff H., Sjoberg M., Cheng R. H. 2004. Budding of alphaviruses. Virus Res. 106:103–116 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
