

A requirement for nuclear factor-kappaB in developmental and plasticity-associated synaptogenesis
- PMID: 21471377
- PMCID: PMC3113725
- DOI: 10.1523/JNEUROSCI.2456-10.2011
A requirement for nuclear factor-kappaB in developmental and plasticity-associated synaptogenesis
Abstract
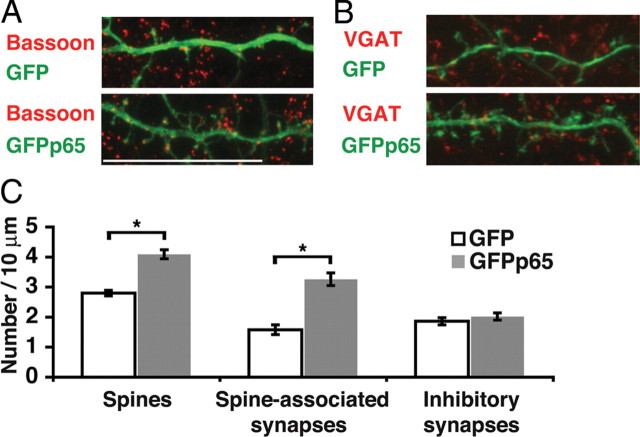
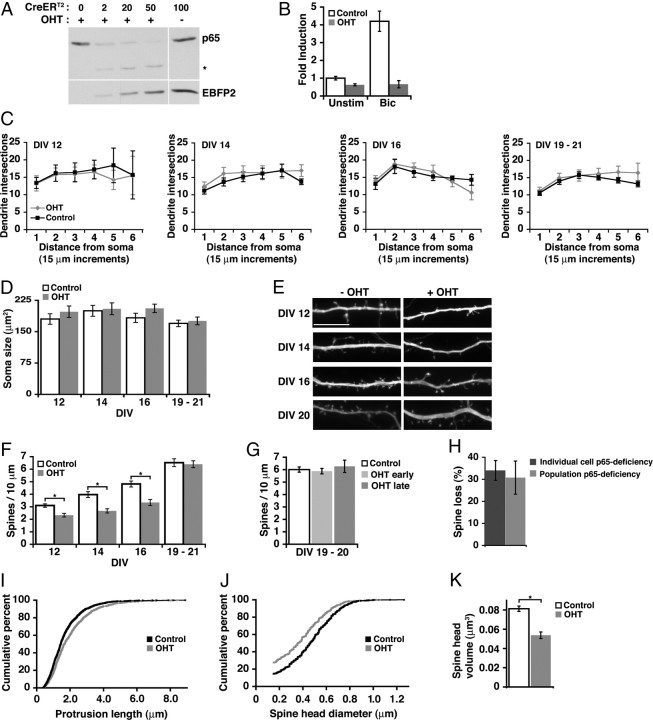
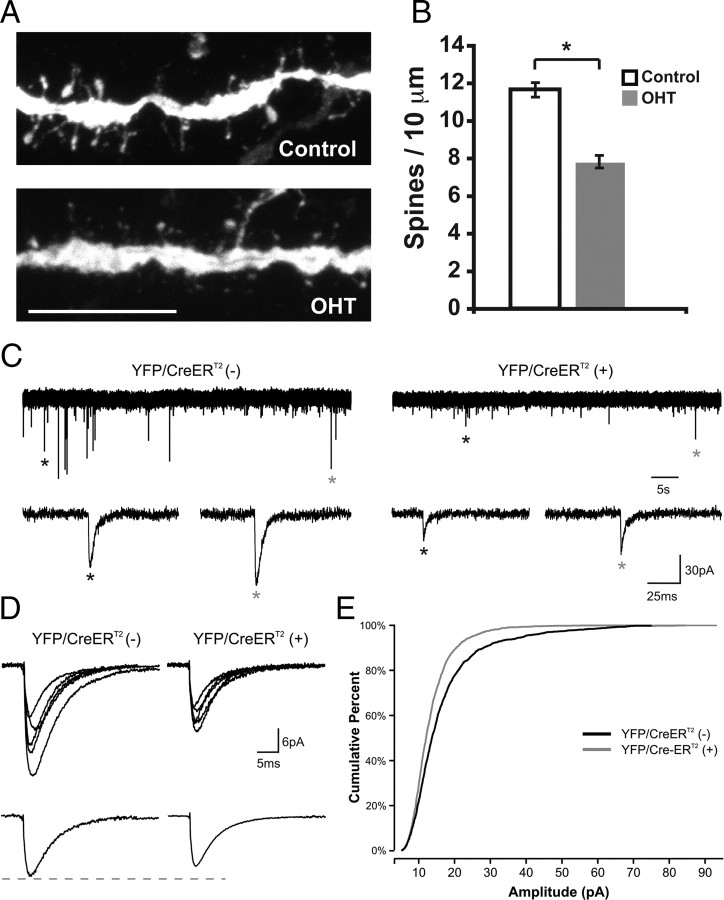
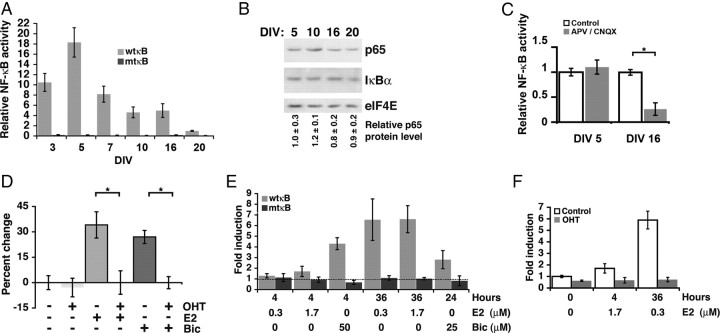
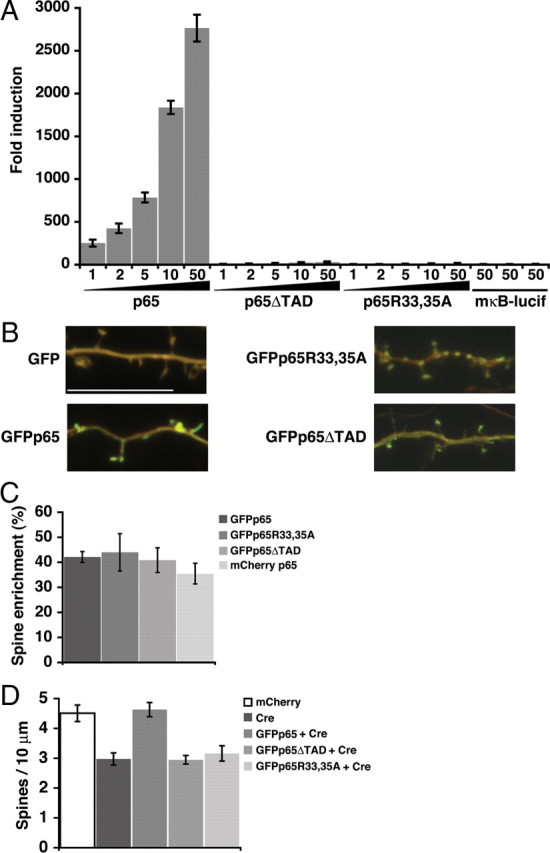
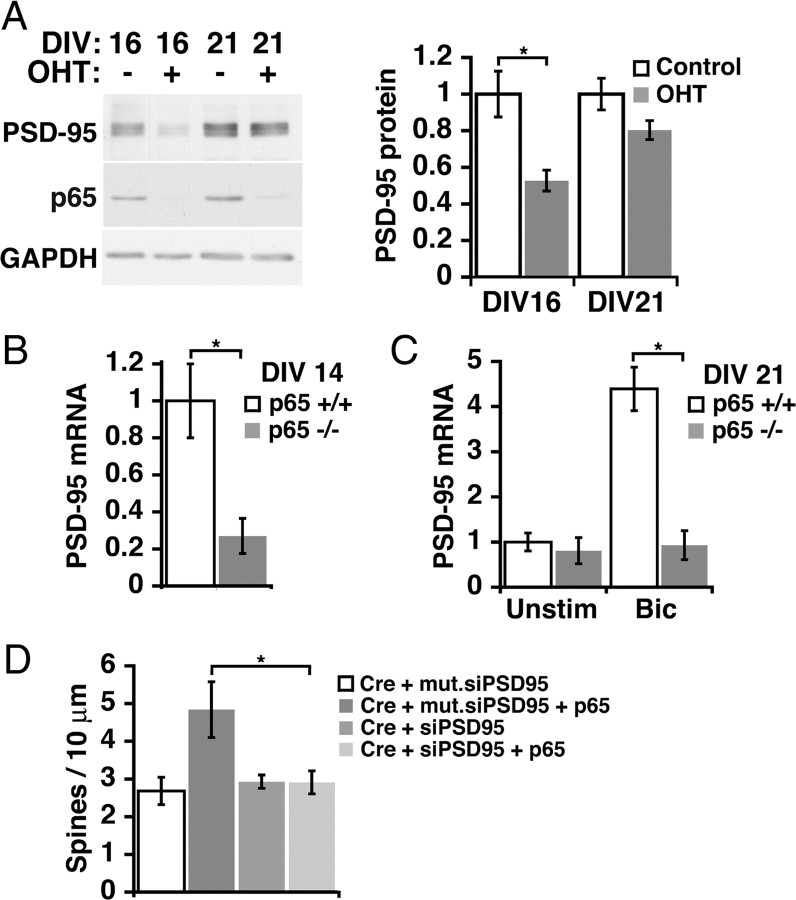
Structural plasticity of dendritic spines and synapses is a fundamental mechanism governing neuronal circuits and may form an enduring basis for information storage in the brain. We find that the p65 subunit of the nuclear factor-κB (NF-κB) transcription factor, which is required for learning and memory, controls excitatory synapse and dendritic spine formation and morphology in murine hippocampal neurons. Endogenous NF-κB activity is elevated by excitatory transmission during periods of rapid spine and synapse development. During in vitro synaptogenesis, NF-κB enhances dendritic spine and excitatory synapse density and loss of endogenous p65 decreases spine density and spine head volume. Cell-autonomous function of NF-κB within the postsynaptic neuron is sufficient to regulate the formation of both presynaptic and postsynaptic elements. During synapse development in vivo, loss of NF-κB similarly reduces spine density and also diminishes the amplitude of synaptic responses. In contrast, after developmental synaptogenesis has plateaued, endogenous NF-κB activity is low and p65 deficiency no longer attenuates basal spine density. Instead, NF-κB in mature neurons is activated by stimuli that induce demand for new synapses, including estrogen and short-term bicuculline, and is essential for upregulating spine density in response to these stimuli. p65 is enriched in dendritic spines making local protein-protein interactions possible; however, the effects of NF-κB on spine density require transcription and the NF-κB-dependent regulation of PSD-95, a critical postsynaptic component. Collectively, our data define a distinct role for NF-κB in imparting transcriptional regulation required for the induction of changes to, but not maintenance of, excitatory synapse and spine density.
Figures
References
-
- Ai HW, Shaner NC, Cheng Z, Tsien RY, Campbell RE. Exploration of new chromophore structures leads to the identification of improved blue fluorescent proteins. Biochemistry. 2007;46:5904–5910. - PubMed
-
- Bhatt DH, Zhang S, Gan WB. Dendritic spine dynamics. Annu Rev Physiol. 2009;71:261–282. - PubMed
-
- Bonhoeffer T, Yuste R. Spine motility. Phenomenology, mechanisms, and function. Neuron. 2002;35:1019–1027. - PubMed
-
- Chen FE, Huang DB, Chen YQ, Ghosh G. Crystal structure of p50/p65 heterodimer of transcription factor NF-kappaB bound to DNA. Nature. 1998;391:410–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases