

Liver autophagy contributes to the maintenance of blood glucose and amino acid levels
- PMID: 21471734
- PMCID: PMC3149698
- DOI: 10.4161/auto.7.7.15371
Liver autophagy contributes to the maintenance of blood glucose and amino acid levels
Abstract
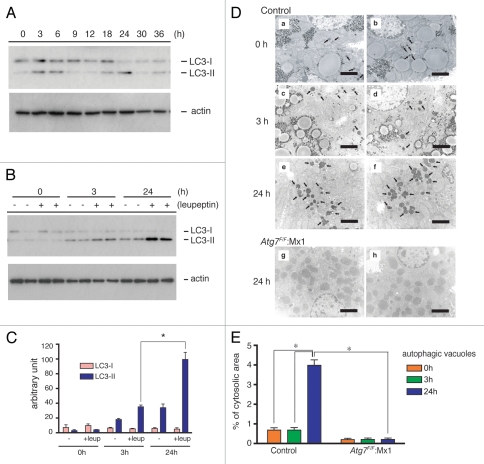
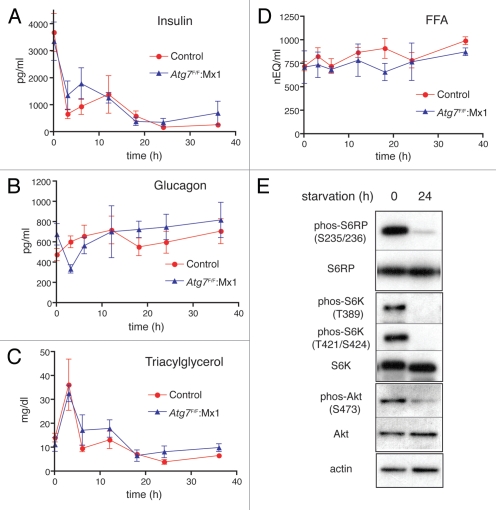
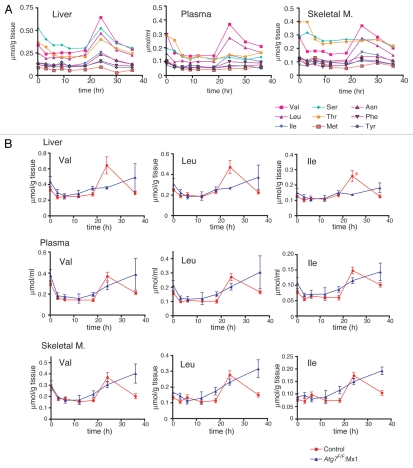
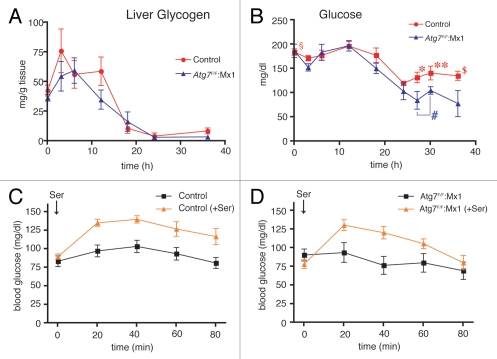
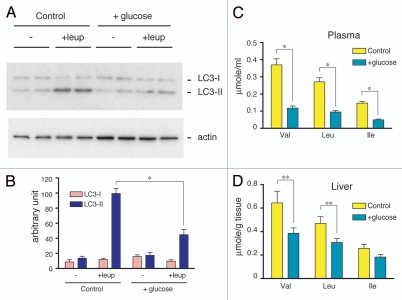
Both anabolism and catabolism of the amino acids released by starvation-induced autophagy are essential for cell survival, but their actual metabolic contributions in adult animals are poorly understood. Herein, we report that, in mice, liver autophagy makes a significant contribution to the maintenance of blood glucose by converting amino acids to glucose via gluconeogenesis. Under a synchronous fasting-initiation regimen, autophagy was induced concomitantly with a fall in plasma insulin in the presence of stable glucagon levels, resulting in a robust amino acid release. In liver-specific autophagy (Atg7)-deficient mice, no amino acid release occurred and blood glucose levels continued to decrease in contrast to those of wild-type mice. Administration of serine (30 mg/animal) exerted a comparable effect, raising the blood glucose levels in both control wild-type and mutant mice under starvation. Thus, the absence of the amino acids that were released by autophagic proteolysis is a major reason for a decrease in blood glucose. Autophagic amino acid release in control wild-type livers was significantly suppressed by the prior administration of glucose, which elicited a prompt increase in plasma insulin levels. This indicates that insulin plays a dominant role over glucagon in controlling liver autophagy. These results are the first to show that liver-specific autophagy plays a role in blood glucose regulation.
Figures
References
-
- Mizushima N. Autophagy: process and function. Genes Dev. 2007;21:2861–2873. - PubMed
-
- Xie Z, Klionsky DJ. Autophagosome formation: core machinery and adaptations. Nat Cell Biol. 2007;9:1102–1109. - PubMed
-
- Amano A, Nakagawa I, Yoshimori T. Autophagy in innate immunity against intracellular bacteria. J Biochem. 2006;140:161–166. - PubMed
-
- Komatsu M, Ueno T, Waguri S, Uchiyama Y, Kominami E, Tanaka K. Constitutive autophagy: vital role in clearance of unfavorable proteins in neurons. Cell Death Differ. 2007;14:887–894. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases