

Dynamic interchanging native states of lymphotactin examined by SNAPP-MS
- PMID: 21472559
- PMCID: PMC3061006
- DOI: 10.1007/s13361-010-0042-3
Dynamic interchanging native states of lymphotactin examined by SNAPP-MS
Abstract
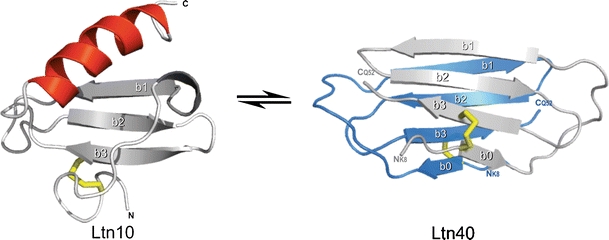
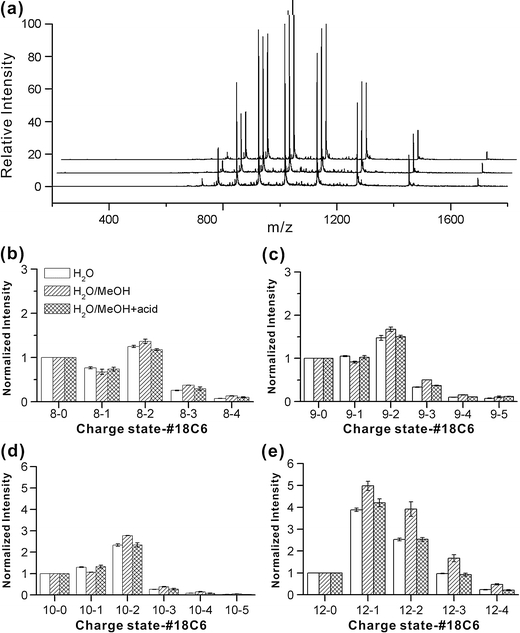
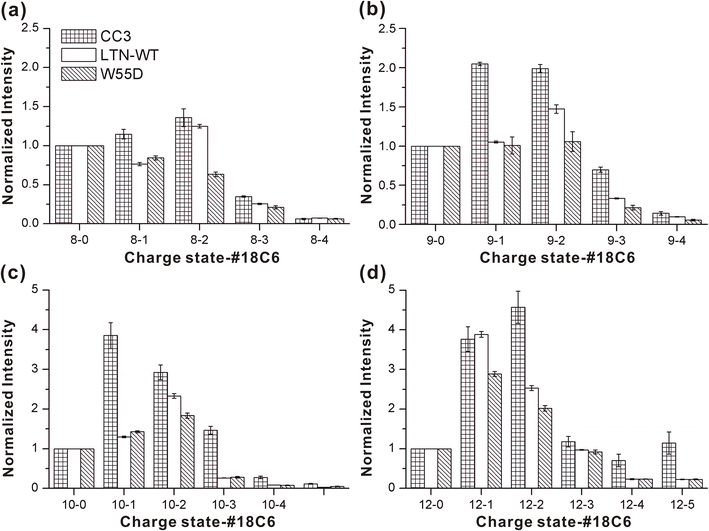
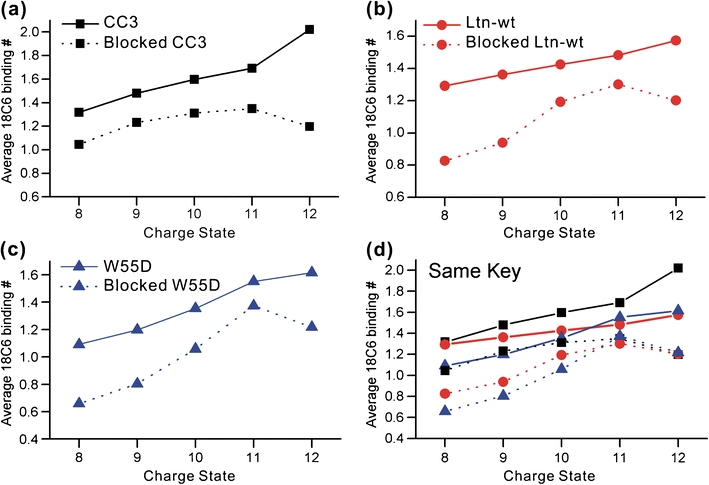
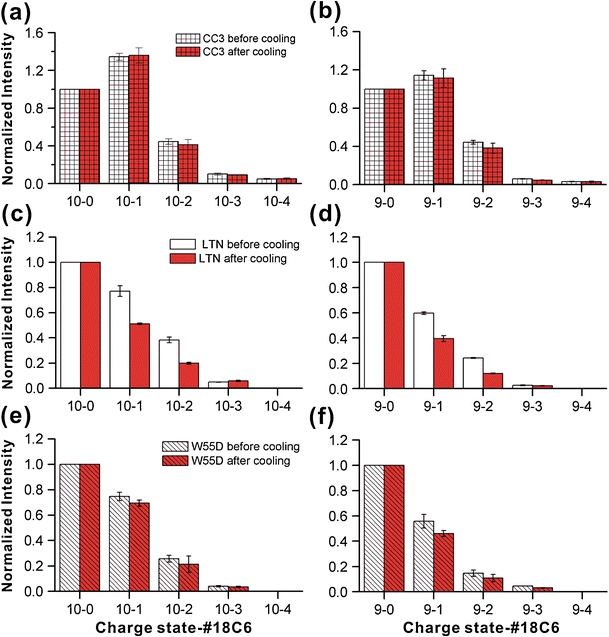
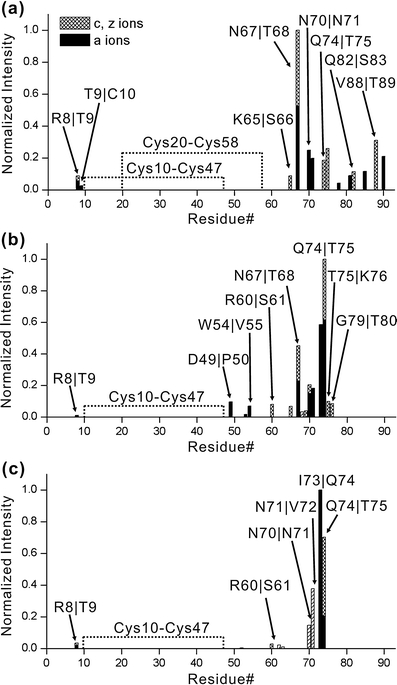
The human chemokine lymphotactin (Ltn) is a remarkable protein that interconverts between two unrelated native state structures in the condensed phase. It is possible to shift the equilibrium toward either conformation with selected sequence substitutions. Previous results have shown that a disulfide-stabilized variant preferentially adopts the canonical chemokine fold (Ltn10), while a single amino acid change (W55D) favors the novel Ltn40 dimeric structure. Selective noncovalent adduct protein probing (SNAPP) is a recently developed method for examining solution phase protein structure. Herein, it is demonstrated that SNAPP can easily recognize and distinguish between the Ltn10 and Ltn40 states of lymphotactin in aqueous solution. The effects of organic denaturants, acid, and disulfide bond reduction and blocking were also examined using SNAPP for the CC3, W55D, and wild type proteins. Only disulfide reduction was shown to significantly perturb the protein, and resulted in considerably decreased adduct formation consistent with loss of tertiary/secondary structure. Cold denaturation experiments demonstrated that wild-type Ltn is the most temperature sensitive of the three proteins. Examination of the higher charge states in all experiments, which are presumed to represent transition state structures between Ltn-10 and Ltn-40, reveals increased 18C6 attachment relative to the more folded structures. This observation is consistent with increased competitive intramolecular hydrogen bonding, which may guide the transition. Experiments examining the gas phase structures revealed that all three proteins can be structurally distinguished in the gas phase. In addition, the gas phase experiments enabled identification of preferred adduct binding sites.
© The Author(s) 2011. This article is published with open access at Springerlink.com
Figures

References
-
- Rodgers, M. T.; Chen, Y. Noncovalent Molecular Recognition of Protonated Peptidomimetic Bases by 18-Crown-6: Structure versus Energetics. Proceedings of the 58th ASMS Conference on Mass Spectrometry and Allied Topics; Salt Lake City, Utah.
