

A single amino acid residue constitutes the third dimerization domain essential for the assembly and function of the tetrameric polycystin-2 (TRPP2) channel
- PMID: 21474446
- PMCID: PMC3099714
- DOI: 10.1074/jbc.M110.192286
A single amino acid residue constitutes the third dimerization domain essential for the assembly and function of the tetrameric polycystin-2 (TRPP2) channel
Abstract
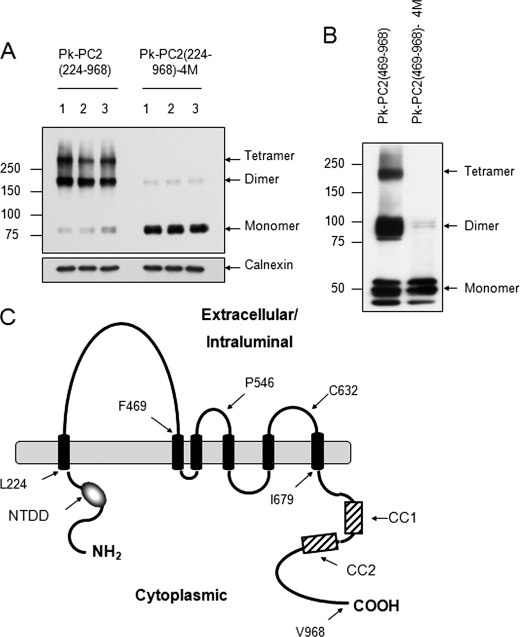
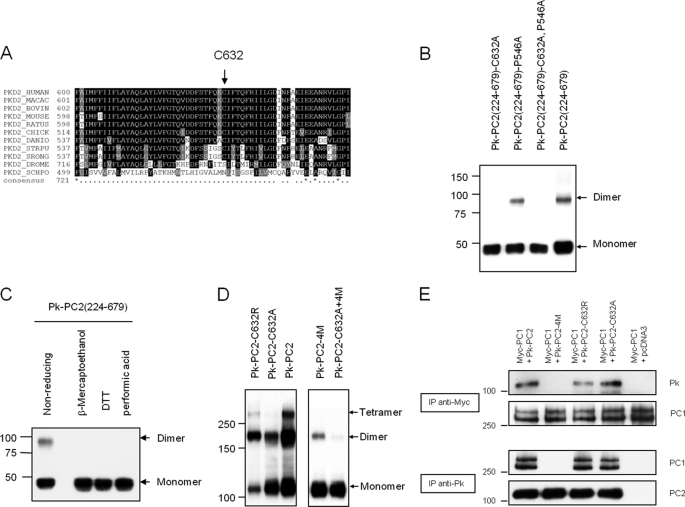
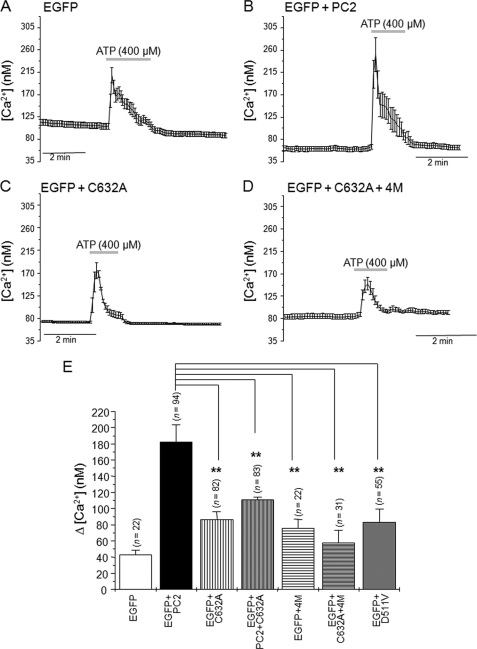
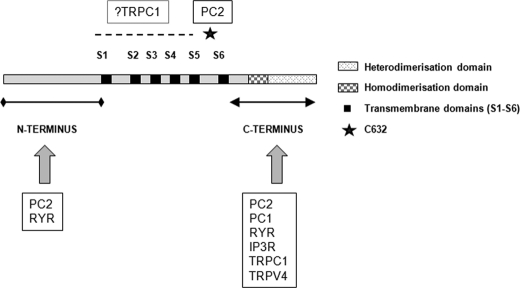
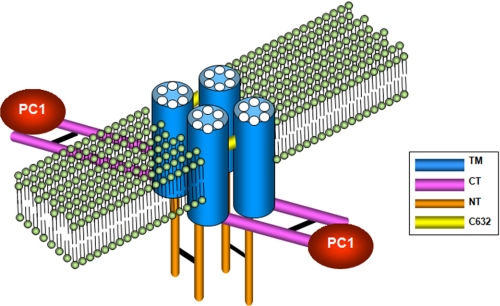
Autosomal dominant polycystic kidney disease (ADPKD), the most common inherited cause of kidney failure, is caused by mutations in either PKD1 (85%) or PKD2 (15%). The PKD2 protein, polycystin-2 (PC2 or TRPP2), is a member of the transient receptor potential (TRP) superfamily and functions as a nonselective calcium channel. PC2 has been found to form oligomers in native tissues, suggesting that similar to other TRP channels, it may form functional homo- or heterotetramers with other TRP subunits. We have recently demonstrated that the homodimerization of PC2 is mediated by both N-terminal and C-terminal domains, and it is known that PC2 can heterodimerize with PC1, TRPC1, and TRPV4. In this paper, we report that a single cysteine residue, Cys(632), mutated in a known PKD2 pedigree, constitutes the third dimerization domain for PC2. PC2 truncation mutants lacking both N and C termini could still dimerize under nonreducing conditions. Mutation of Cys(632) alone abolished dimerization in these mutants, indicating that it was the critical residue mediating disulfide bond formation between PC2 monomers. Co-expression of C632A PC2 mutants with wild-type PC2 channels reduced ATP-sensitive endoplasmic reticulum Ca(2+) release in HEK293 cells. The combination of C632A and mutations disrupting the C-terminal coiled-coil domain (Val(846), Ile(853), Ile(860), Leu(867) or 4M) nearly abolished dimer formation and ATP-dependent Ca(2+) release. However, unlike the 4M PC2 mutant, a C632A mutant could still heterodimerize with polycystin-1 (PC1). Our results indicate that PC2 homodimerization is regulated by three distinct domains and that these events regulate formation of the tetrameric PC2 channel.
Figures
References
-
- Calvet J. P., Grantham J. J. (2001) Semin. Nephrol. 21, 107–123 - PubMed
-
- Wilson P. D. (2004) N. Engl. J. Med. 350, 151–164 - PubMed
-
- Mochizuki T., Wu G., Hayashi T., Xenophontos S. L., Veldhuisen B., Saris J. J., Reynolds D. M., Cai Y., Gabow P. A., Pierides A., Kimberling W. J., Breuning M. H., Deltas C. C., Peters D. J., Somlo S. (1996) Science 272, 1339–1342 - PubMed
-
- Montell C., Birnbaumer L., Flockerzi V., Bindels R. J., Bruford E. A., Caterina M. J., Clapham D. E., Harteneck C., Heller S., Julius D., Kojima I., Mori Y., Penner R., Prawitt D., Scharenberg A. M., Schultz G., Shimizu N., Zhu M. X. (2002) Mol. Cell 9, 229–231 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
