

Integrin β7-mediated regulation of multiple myeloma cell adhesion, migration, and invasion
- PMID: 21474670
- PMCID: PMC3122944
- DOI: 10.1182/blood-2010-06-292243
Integrin β7-mediated regulation of multiple myeloma cell adhesion, migration, and invasion
Erratum in
-
Neri P, Ren L, Azab AK, et al. Integrin β7-mediated regulation of multiple myeloma cell adhesion, migration, and invasion. Blood. 2011;117(23):6202-6213.Blood. 2025 Nov 20;146(21):2613. doi: 10.1182/blood.2025031937. Blood. 2025. PMID: 41264322 Free PMC article. No abstract available.
Abstract
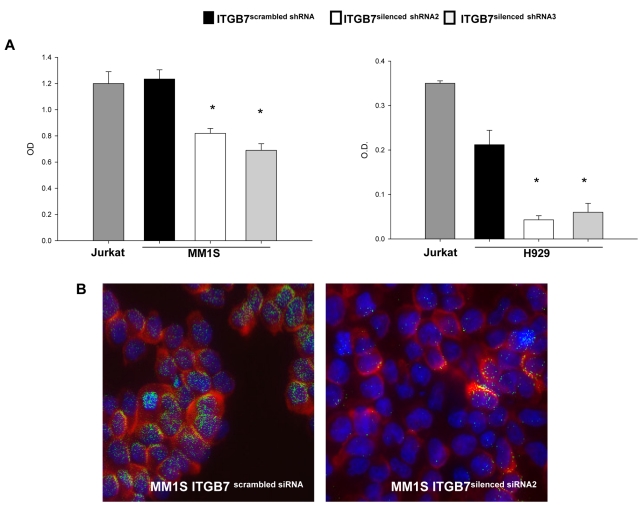
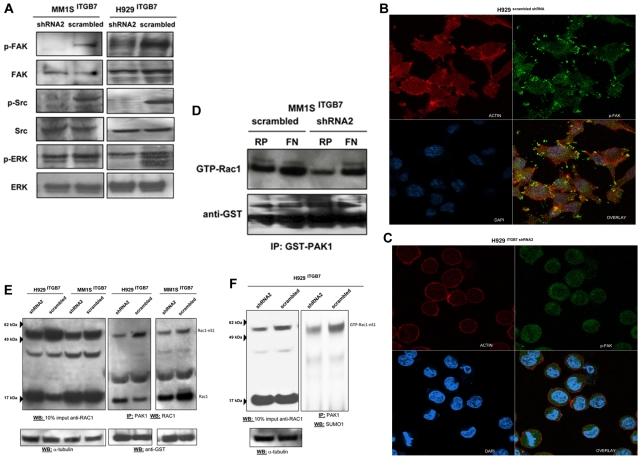
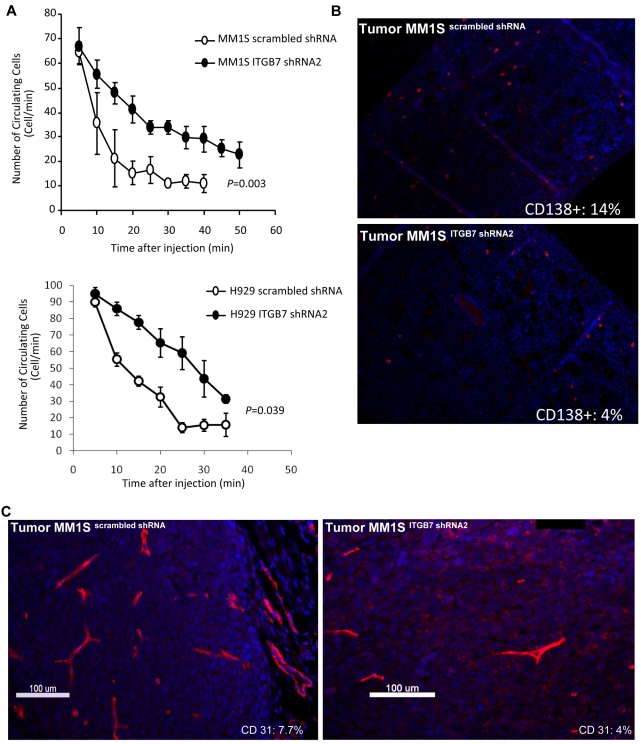
Integrin-β7 (ITGB7) mRNA is detected in multiple myeloma (MM) cells and its presence is correlated with MAF gene activation. Although the involvement of several integrin family members in MM-stoma cell interaction is well documented, the specific biologic functions regulated by integrin-β7 in MM are largely unknown. Clinically, we have correlated integrin-β7 expression in MM with poor survival outcomes post autologous stem cell transplantation and postsalvage therapy with bortezomib. Functionally, we have found that shRNA-mediated silencing of ITGB7 reduces MM-cell adhesion to extra-cellular matrix elements (fibronectin, E-cadherin) and reverses cell-adhesion-mediated drug resistance (CAM-DR) sensitizing them to bortezomib and melphalan. In addition, ITGB7 silencing abrogated MM-cell transwell migration in response to SDF1α gradients, reduced vessel density in xenografted tumors, and altered MM cells in vivo homing into the BM. Mechanistically, ITGB7 knockdown inhibited focal adhesion kinase (FAK) and Src phosphorylation, Rac1 activation, and SUMOylation, reduced VEGF production in MM-BM stem cell cocultures and attenuated p65-NF-κB activity. Our findings support a role for integrin-β7 in MM-cell adhesion, migration, and BM homing, and pave the way for a novel therapeutic approach targeting this molecule.
Figures
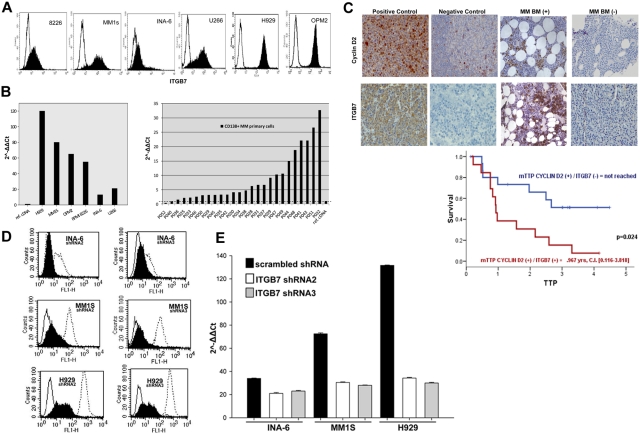
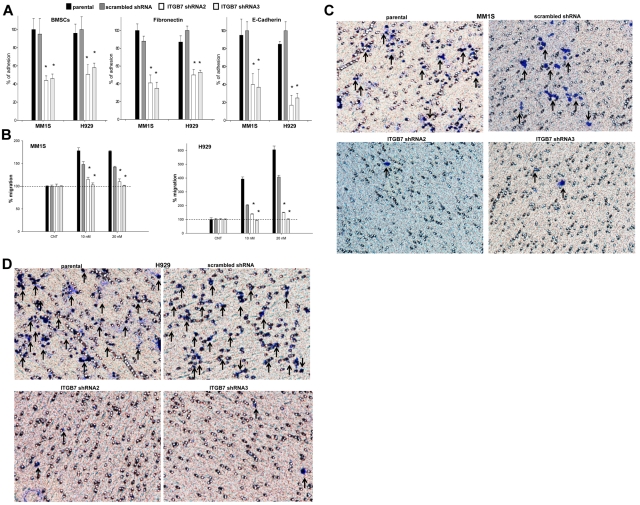
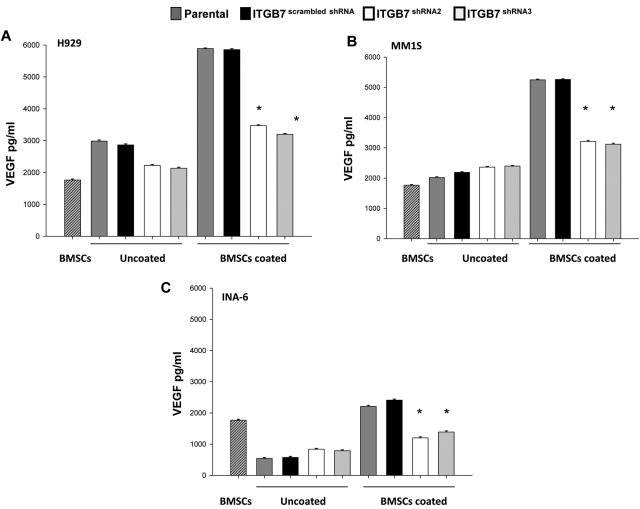
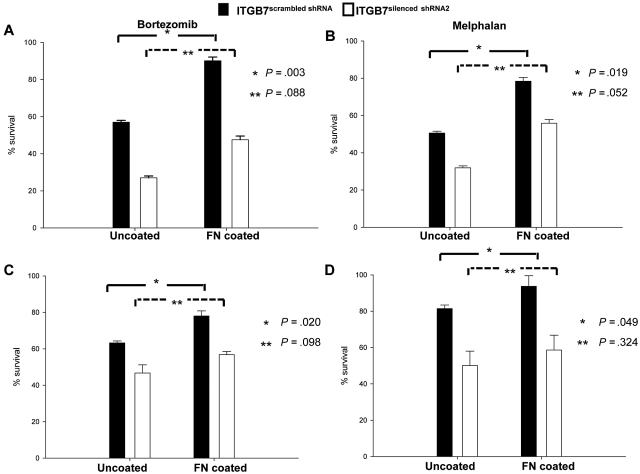
References
-
- Richardson PG, Sonneveld P, Schuster MW, et al. Bortezomib or high-dose dexamethasone for relapsed multiple myeloma. N Engl J Med. 2005;352(24):2487–2498. - PubMed
-
- Dimopoulos M, Spencer A, Attal M, et al. Lenalidomide plus dexamethasone for relapsed or refractory multiple myeloma. N Engl J Med. 2007;357(21):2123–2132. - PubMed
-
- Chauhan D, Uchiyama H, Akbarali Y, et al. Multiple myeloma cell adhesion-induced interleukin-6 expression in bone marrow stromal cells involves activation of NF-kappa B. Blood. 1996;87(3):1104–1112. - PubMed
-
- Podar K, Tai YT, Davies FE, et al. Vascular endothelial growth factor triggers signaling cascades mediating multiple myeloma cell growth and migration. Blood. 2001;98(2):428–435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
