

Ataxin-1 and Brother of ataxin-1 are components of the Notch signalling pathway
- PMID: 21475249
- PMCID: PMC3090018
- DOI: 10.1038/embor.2011.49
Ataxin-1 and Brother of ataxin-1 are components of the Notch signalling pathway
Abstract
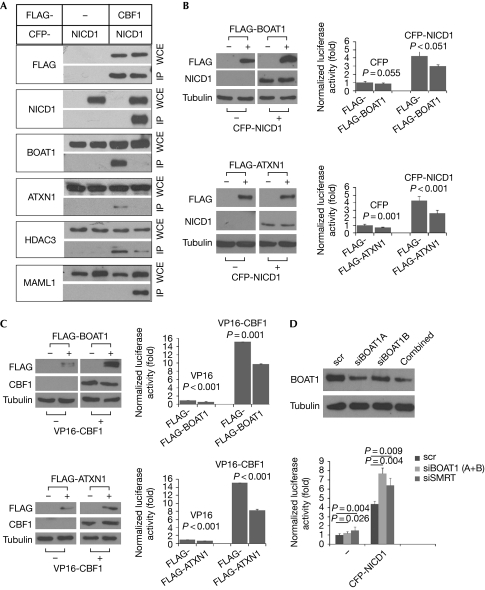
Ataxin-1 (ATXN1), a causative factor for spinocerebellar ataxia type 1 (SCA1), and the related Brother of ATXN1 (BOAT1) are human proteins involved in transcriptional repression. So far, little is known about which transcriptional pathways mediate the effects of ATXN1 and BOAT1. From our analyses of the properties of BOAT1 in Drosophila and of both proteins in mammalian cells, we report here that BOAT1 and ATXN1 are components of the Notch signalling pathway. In Drosophila, BOAT1 compromises the activities of Notch. In mammalian cells, both ATXN1 and BOAT1 bind to the promoter region of Hey1 and inhibit the transcriptional output of Notch through direct interactions with CBF1, a transcription factor that is crucial for the Notch pathway. Our results suggest that, in addition to their involvement in SCA1, ATXN1 and BOAT1 might participate in several Notch-controlled developmental and pathological processes.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



Similar articles
-
The role of LANP and ataxin 1 in E4F-mediated transcriptional repression.EMBO Rep. 2007 Jul;8(7):671-7. doi: 10.1038/sj.embor.7400983. Epub 2007 Jun 8. EMBO Rep. 2007. PMID: 17557114 Free PMC article.
-
Structural basis of protein complex formation and reconfiguration by polyglutamine disease protein Ataxin-1 and Capicua.Genes Dev. 2013 Mar 15;27(6):590-5. doi: 10.1101/gad.212068.112. Genes Dev. 2013. PMID: 23512657 Free PMC article.
-
Partial loss of ataxin-1 function contributes to transcriptional dysregulation in spinocerebellar ataxia type 1 pathogenesis.PLoS Genet. 2010 Jul 8;6(7):e1001021. doi: 10.1371/journal.pgen.1001021. PLoS Genet. 2010. PMID: 20628574 Free PMC article.
-
Hairless: the ignored antagonist of the Notch signalling pathway.Hereditas. 2006 Dec;143(2006):212-21. doi: 10.1111/j.2007.0018-0661.01971.x. Hereditas. 2006. PMID: 17362357 Review.
-
Notch-independent functions of CSL.Curr Top Dev Biol. 2011;97:55-74. doi: 10.1016/B978-0-12-385975-4.00009-7. Curr Top Dev Biol. 2011. PMID: 22074602 Review.
Cited by
-
Hsa_circ_0008309 May Be a Potential Biomarker for Oral Squamous Cell Carcinoma.Dis Markers. 2018 Sep 23;2018:7496890. doi: 10.1155/2018/7496890. eCollection 2018. Dis Markers. 2018. PMID: 30344795 Free PMC article.
-
Circular RNAs and their roles in head and neck cancers.Mol Cancer. 2019 Mar 21;18(1):44. doi: 10.1186/s12943-019-1003-5. Mol Cancer. 2019. PMID: 30898135 Free PMC article. Review.
-
The expanding role for chromatin and transcription in polyglutamine disease.Curr Opin Genet Dev. 2014 Jun;26:96-104. doi: 10.1016/j.gde.2014.06.008. Epub 2014 Aug 11. Curr Opin Genet Dev. 2014. PMID: 25108806 Free PMC article. Review.
-
Integrated molecular analysis of adult T cell leukemia/lymphoma.Nat Genet. 2015 Nov;47(11):1304-15. doi: 10.1038/ng.3415. Epub 2015 Oct 5. Nat Genet. 2015. PMID: 26437031
-
The vasculome of the mouse brain.PLoS One. 2012;7(12):e52665. doi: 10.1371/journal.pone.0052665. Epub 2012 Dec 20. PLoS One. 2012. PMID: 23285140 Free PMC article.
References
-
- Banfi S, Servadio A, Chung MY, Kwiatkowski TJ Jr, McCall AE, Duvick LA, Shen Y, Roth EJ, Orr HT, Zoghbi HY (1994) Identification and characterization of the gene causing type 1 spinocerebellar ataxia. Nat Genet 7: 513–520 - PubMed
-
- Bolger TA, Zhao X, Cohen TJ, Tsai CC, Yao TP (2007) The neurodegenerative disease protein ataxin-1 antagonizes the neuronal survival function of myocyte enhancer factor-2. J Biol Chem 282: 29186–29192 - PubMed
-
- Bray SJ (2006) Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol 7: 678–689 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
