

Multilevel control of Arabidopsis 3-hydroxy-3-methylglutaryl coenzyme A reductase by protein phosphatase 2A
- PMID: 21478440
- PMCID: PMC3101556
- DOI: 10.1105/tpc.110.074278
Multilevel control of Arabidopsis 3-hydroxy-3-methylglutaryl coenzyme A reductase by protein phosphatase 2A
Abstract
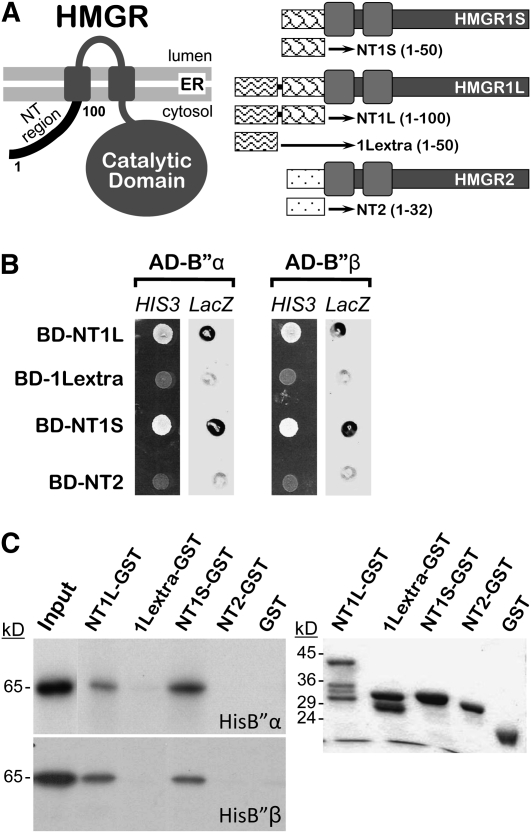
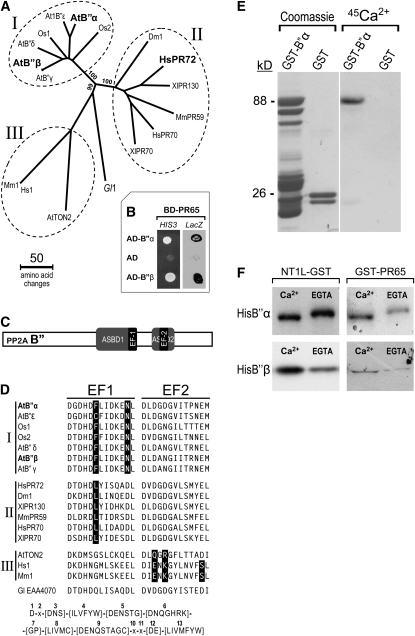
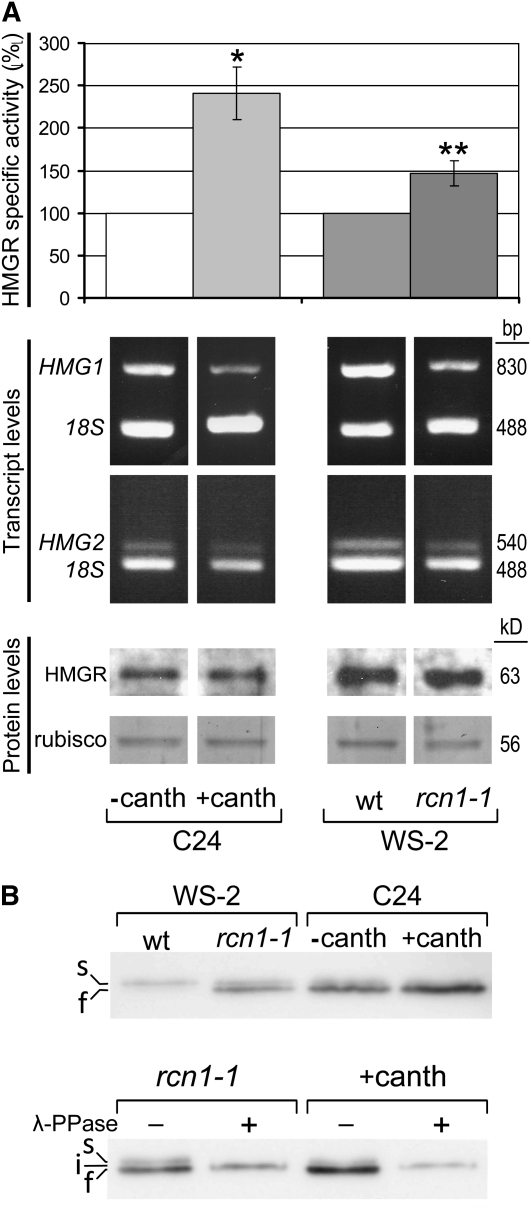
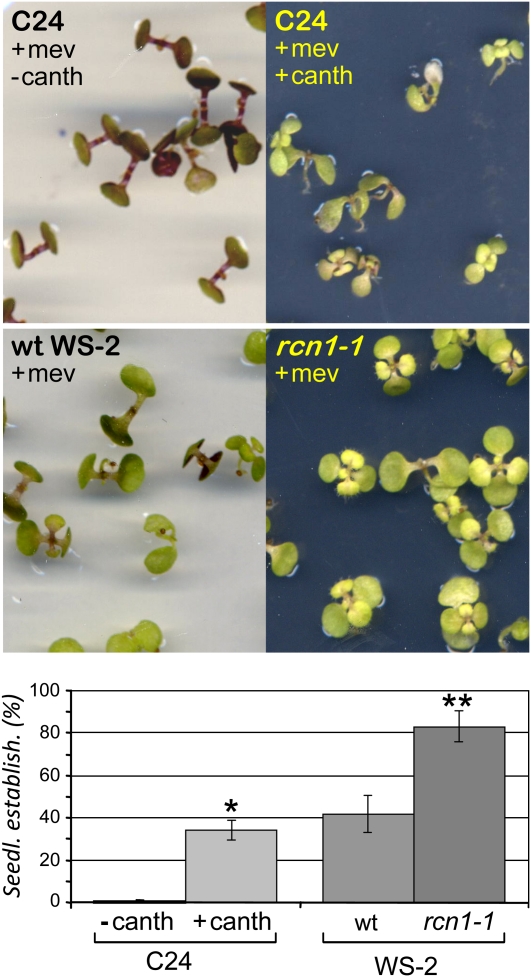
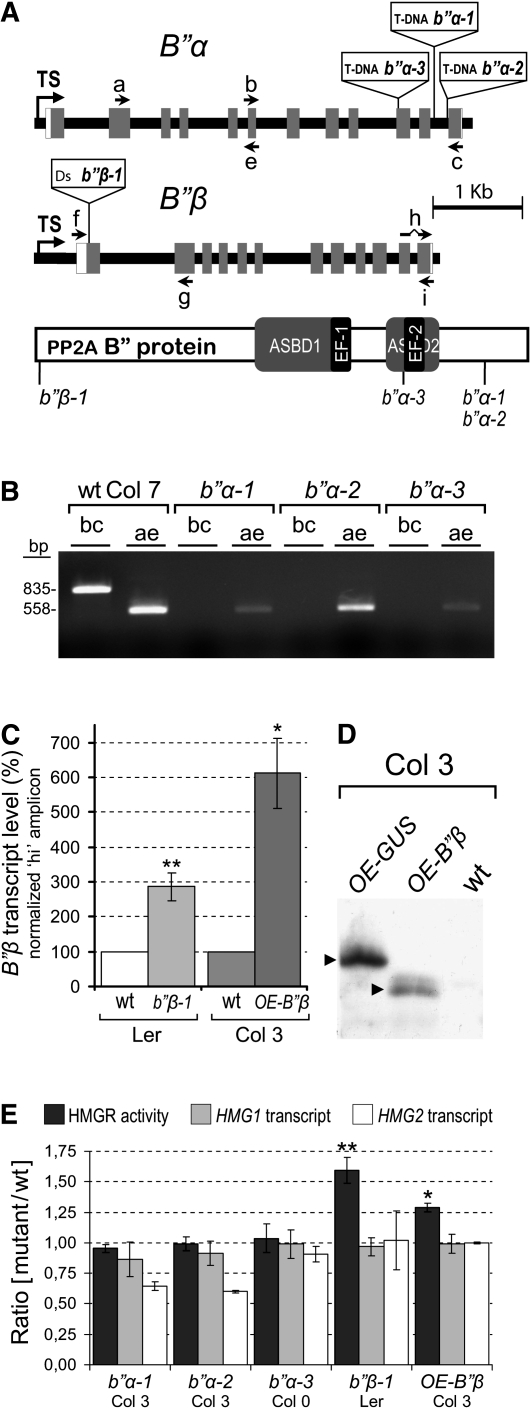
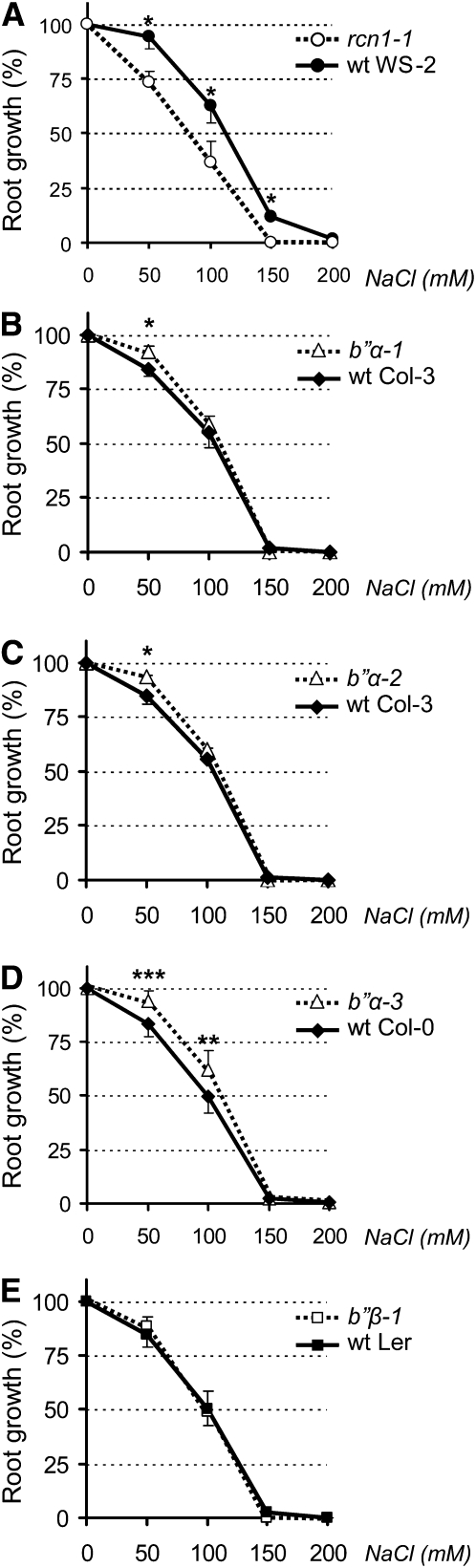
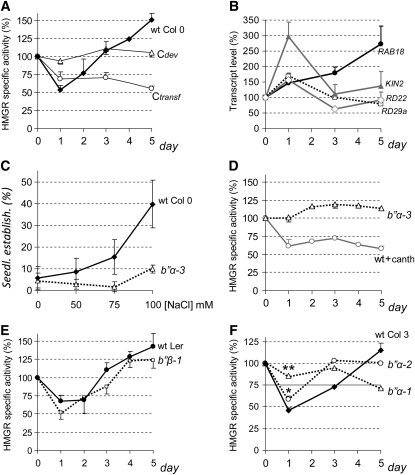
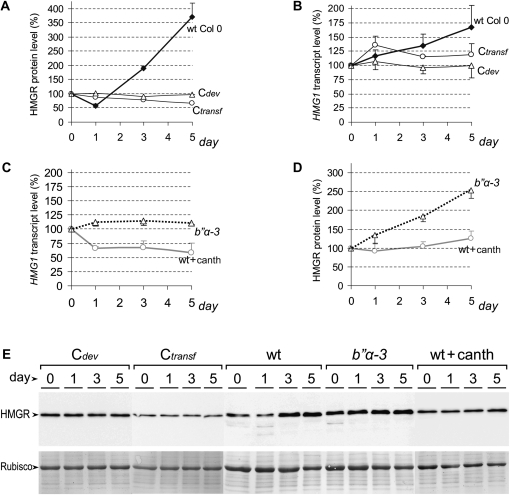
Plants synthesize a myriad of isoprenoid products that are required both for essential constitutive processes and for adaptive responses to the environment. The enzyme 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR) catalyzes a key regulatory step of the mevalonate pathway for isoprenoid biosynthesis and is modulated by many endogenous and external stimuli. In spite of that, no protein factor interacting with and regulating plant HMGR in vivo has been described so far. Here, we report the identification of two B'' regulatory subunits of protein phosphatase 2A (PP2A), designated B''α and B''β, that interact with HMGR1S and HMGR1L, the major isoforms of Arabidopsis thaliana HMGR. B''α and B''β are Ca²⁺ binding proteins of the EF-hand type. We show that HMGR transcript, protein, and activity levels are modulated by PP2A in Arabidopsis. When seedlings are transferred to salt-containing medium, B''α and PP2A mediate the decrease and subsequent increase of HMGR activity, which results from a steady rise of HMGR1-encoding transcript levels and an initial sharper reduction of HMGR protein level. In unchallenged plants, PP2A is a posttranslational negative regulator of HMGR activity with the participation of B''β. Our data indicate that PP2A exerts multilevel control on HMGR through the five-member B'' protein family during normal development and in response to a variety of stress conditions.
Figures
References
-
- Ausubel F.M., Brent R., Kingston R.E., Moore D.D., Seidman J.G., Smith J.A., Struhl K. (1987). Current Protocols in Molecular Biology. (New York: John Wiley and Sons; ).
-
- Bach T.J., Rogers D.H., Rudney H. (1986). Detergent-solubilization, purification, and characterization of membrane-bound 3-hydroxy-3-methylglutaryl-coenzyme A reductase from radish seedlings. Eur. J. Biochem. 154: 103–111 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
