

doi: 10.1534/genetics.111.127191.
Proximodistal patterning in the Drosophila leg: models and mutations
Affiliations
- PMID: 21478472
- PMCID: PMC3070510
- DOI: 10.1534/genetics.111.127191
Item in Clipboard
Proximodistal patterning in the Drosophila leg: models and mutations
Genetics.
2011 Apr.
Abstract
Limbs have a proximodistal axis that usually is not apparent early in development, a striking example of epigenesis. The proximodistal axis was the subject of experimental and theoretical study before any molecular genetic understanding emerged. As developmental genetic studies in Drosophila advanced, the descriptive polar coordinate model of the 1970s evolved into an understanding of how preexisting developmental compartments interact to express signaling molecules, including Hedgehog, Wingless, and Decapentaplegic, and how these define a proximodistal axis as limbs appear.
Figures
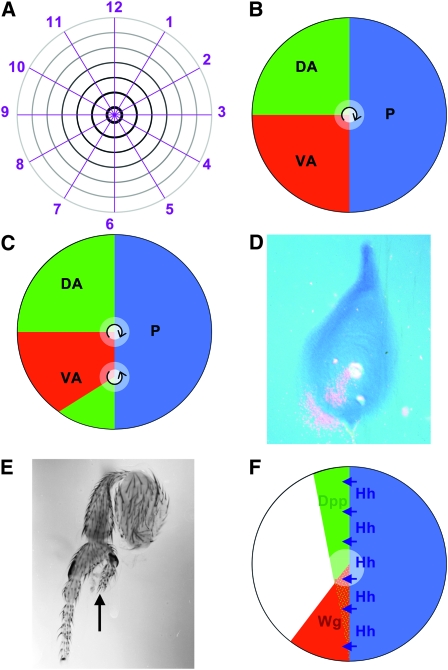
(A) The polar coordinate model represented the surface of limbs and appendages collapsed onto a two-dimensional disc, as is actually the case for Drosophila imaginal discs before they evert during pupation (French
et al. 1976). Cells are thought to have circumferential information, represented by numbers 1–12 as in a clockface, and proximodistal information radiating form the distal center (black) to the proximal periphery (gray). Circumferential regulation is thought to occur by intercalation by the shortest route. Thus, removing less than half the values, e.g., 2–6, permits regeneration of the missing values, but removing more than half the values, e.g., 2–9, results in a duplication of the remaining values 10–1. Missing distal values are generated wherever the entire clockface 1–12 is present, such as in the center of the normal limb (French
et al. 1976). (B) The boundary model posited non-autonomous signals emanating from at least three zones, such as the ventral-anterior (red), dorsal-anterior (green), and posterior (blue). Cells would measure circumferential location, and polarity (arrow) by comparing the red, green, and blue signals. Peak levels of all three signals define the distal point (white), which could be the source of a proximodistal morphogen (Meinhardt 1983). (C) In the boundary model, respecification of patches of cells could promote limb duplications and triplications of various polarities. C shows how respecification of a patch of ventral-anterior cells as dorsal-anterior could lead to a second proximodistal axis with reverse polarity. Respecified patches in other locations could give rise to other defects, depending on the number of points at which all three zones intersect: smaller respecified patches that intersect the anterior-posterior boundary on both sides could give rise to triplications; patches closer to the normal distal tip could alter circumferential patterns without much proximodistal effect. These possibilities are discussed in much more detail by Meinhardt (1983), although he used respecification of posterior cells as the example. Here the example is ventral-anterior because of the correspondence with wingless. (D) Wingless expression in a third instar leg imaginal disc, detected using in situ hybridization with a radioactive probe complementary to the wingless transcript, the main method available in the 1980s. Autoradiography of the dried, hybridized tissue reveals signal as silver grains, shown in pink superimposed over a phase-contrast image of the tissue. Third instar wg expression defines a ventral-anterior domain similar to that hypothesized in the boundary model (Baker 1988b). (E) Foreleg dissected from a hypomorphic wg mutant genotype (wgCX3/wgCX4) (Baker 1988b). The duplicated proximodistal axis (arrow) has reversed polarity and the line of mirror symmetry, which extends basally to the femur, runs through ventral-anterior pattern elements, exactly as the Boundary Model predicted for patchy loss of ventral-anterior identity (Figure 1C). Legs from wg mutants show a range of pattern defects that could be explained by defects at different locations within the ventral-anterior domain (Meinhardt 1983; Baker 1988b). The effect on gene transcription of wgCX3 and wgP, two alleles with similar phenotypes and breakpoints at the 3′ end of the wg gene (Baker 1987), have not been studied, but other studies of mutual antagonism between wg and Dpp expression suggests that ventral cells losing wg would adopt dorsal fate and express Dpp (Brook and Cohen 1996). (F) Unlike Hh expression, which is stably conferred by inheritance of the posterior compartment selector gene from embryogenesis, expression of Dpp and wg is induced close to the posterior compartment by Hh (Basler and Struhl 1994). Expression of wg is limited to ventral cells where it limits the expression and function of Dpp. High levels of Wg and Dpp signaling in the center of the leg disc confer distal fates on cells in this location. The most anterior cells (unlabeled) remain proximal because their low exposure to Hh indirectly reduces levels of Wg and Dpp signaling (Basler and Struhl 1994). Although Wg and Dpp signaling must collaborate in the expression of distal genes (Kim
et al. 1996; Lecuit and Cohen 1997), they antagonize one another in dorsal and ventral patterning (Struhl and Basler 1993; Brook and Cohen 1996).
References
-
- Abu-Shaar, M., and R. S. Mann, 1998. Generation of multiple antagonistic domains along the proximodistal axis during Drosophila leg development. Development 125 3821–3830. - PubMed
-
- Anderson, D. T., 1963. The embryology of Dacus tryoni. 2. Development of imaginal discs. J. Embryol. Exp. Morphol. 11 339–351. - PubMed
-
- Ashburner, M. A., 1993. Epilogue, pp. 1493–1506 in The Development of Drosophila melanogaster, edited by M. Bate and A. Martinez-Arias. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Baker, N. E., 1988. a Embryonic and imaginal requirements for wingless, a segment polarity gene in Drosophila. Dev. Biol. 125 96–108. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
