

Functional specificity of local synaptic connections in neocortical networks
- PMID: 21478872
- PMCID: PMC3089591
- DOI: 10.1038/nature09880
Functional specificity of local synaptic connections in neocortical networks
Abstract
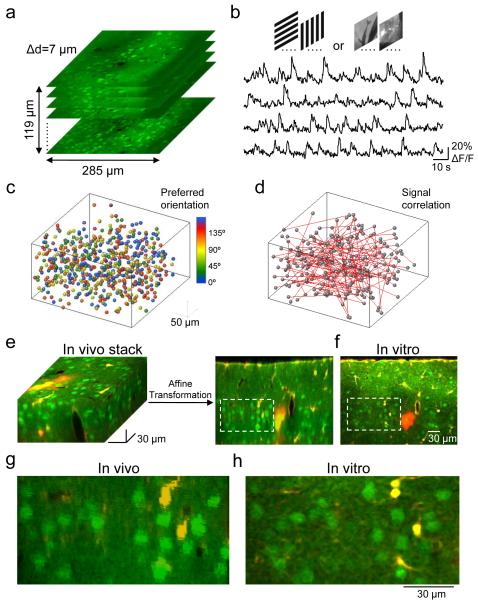
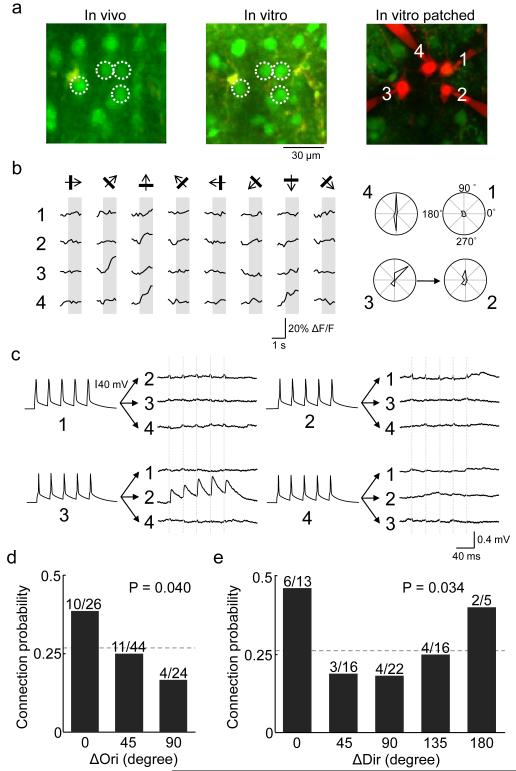
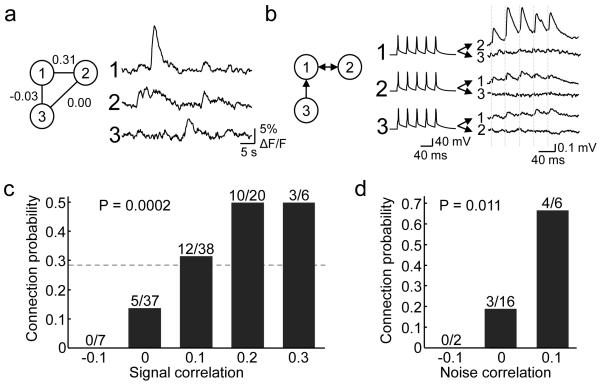
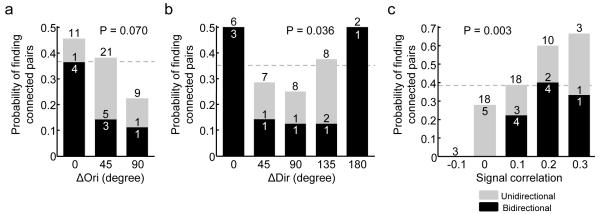
Neuronal connectivity is fundamental to information processing in the brain. Therefore, understanding the mechanisms of sensory processing requires uncovering how connection patterns between neurons relate to their function. On a coarse scale, long-range projections can preferentially link cortical regions with similar responses to sensory stimuli. But on the local scale, where dendrites and axons overlap substantially, the functional specificity of connections remains unknown. Here we determine synaptic connectivity between nearby layer 2/3 pyramidal neurons in vitro, the response properties of which were first characterized in mouse visual cortex in vivo. We found that connection probability was related to the similarity of visually driven neuronal activity. Neurons with the same preference for oriented stimuli connected at twice the rate of neurons with orthogonal orientation preferences. Neurons responding similarly to naturalistic stimuli formed connections at much higher rates than those with uncorrelated responses. Bidirectional synaptic connections were found more frequently between neuronal pairs with strongly correlated visual responses. Our results reveal the degree of functional specificity of local synaptic connections in the visual cortex, and point to the existence of fine-scale subnetworks dedicated to processing related sensory information.
©2011 Macmillan Publishers Limited. All rights reserved
Figures
Comment in
-
Neuronal networks: networking in the visual cortex.Nat Rev Neurosci. 2011 Jun;12(6):306. doi: 10.1038/nrn3041. Epub 2011 Apr 29. Nat Rev Neurosci. 2011. PMID: 21527950 No abstract available.
References
References Methods
-
- Smith SL, Trachtenberg JT. Experience-dependent binocular competition in the visual cortex begins at eye opening. Nat. Neurosci. 2007;10:370–375. - PubMed
-
- Brainard DH. The Psychophysics Toolbox. Spat Vis. 1997;10:433–436. - PubMed
-
- Pelli DG. The VideoToolbox software for visual psychophysics: transforming numbers into movies. Spat Vis. 1997;10:437–442. - PubMed
References
-
- Weliky M, Kandler K, Fitzpatrick D, Katz LC. Patterns of excitation and inhibition evoked by horizontal connections in visual cortex share a common relationship to orientation columns. Neuron. 1995;15:541–552. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
