

A new subtype of progenitor cell in the mouse embryonic neocortex
- PMID: 21478886
- PMCID: PMC3083489
- DOI: 10.1038/nn.2807
A new subtype of progenitor cell in the mouse embryonic neocortex
Abstract
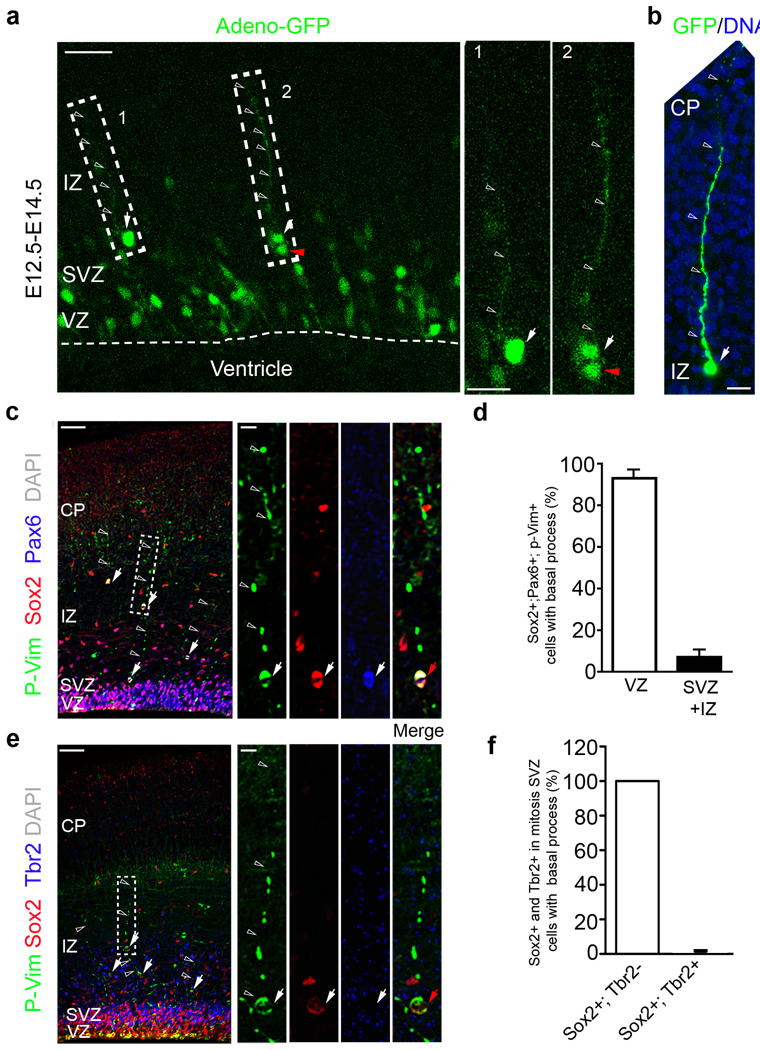
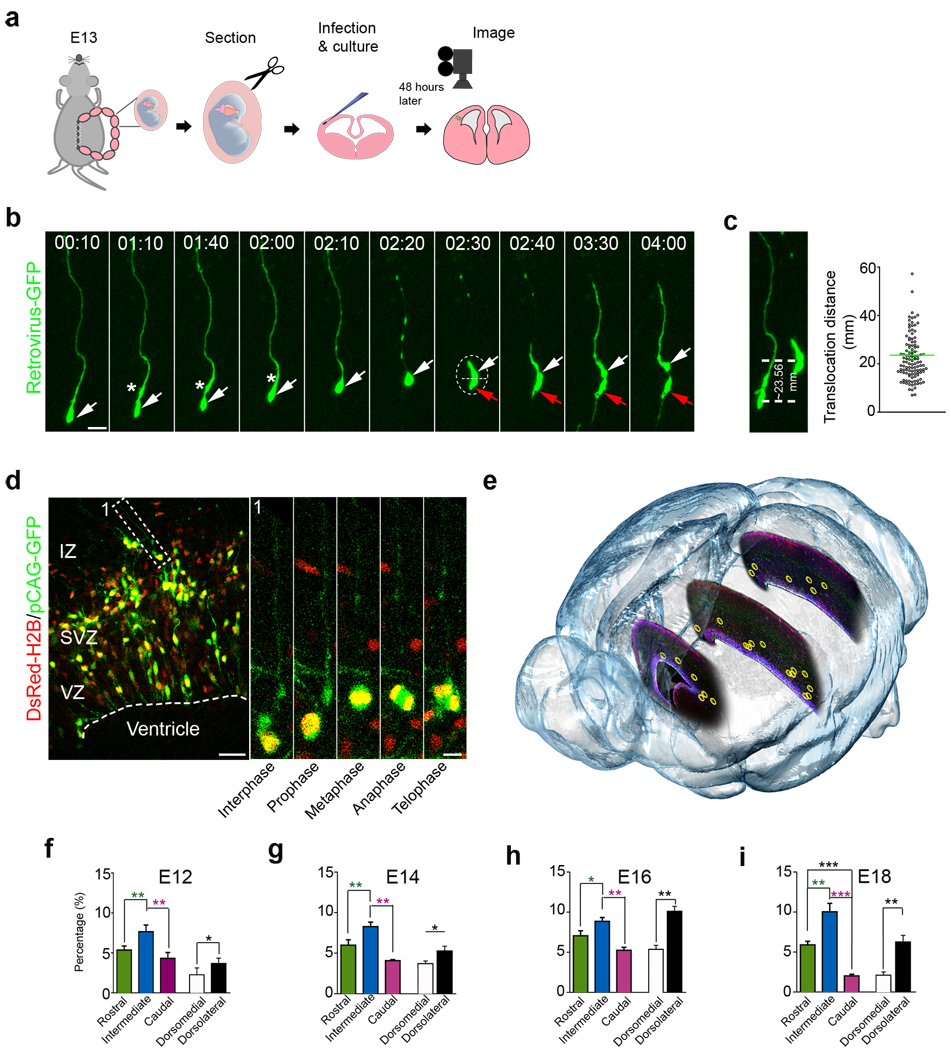
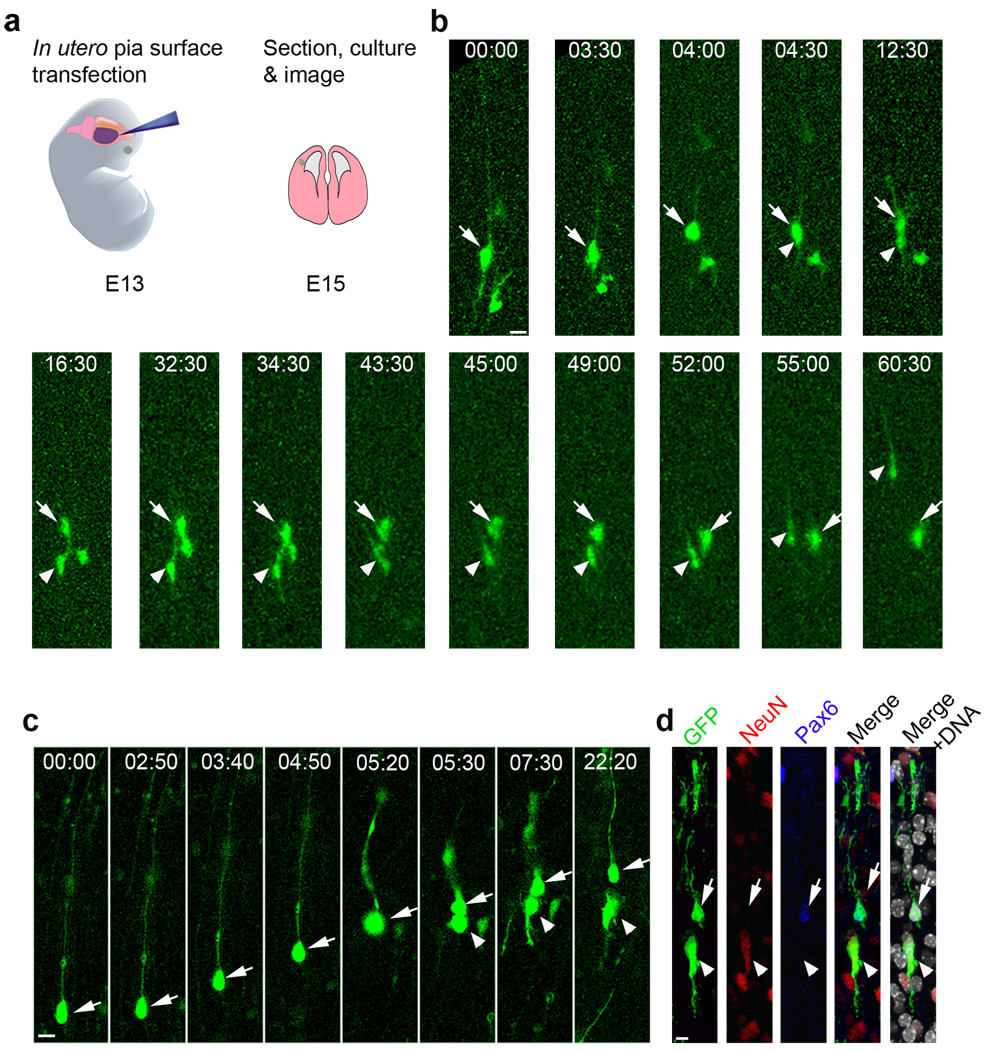
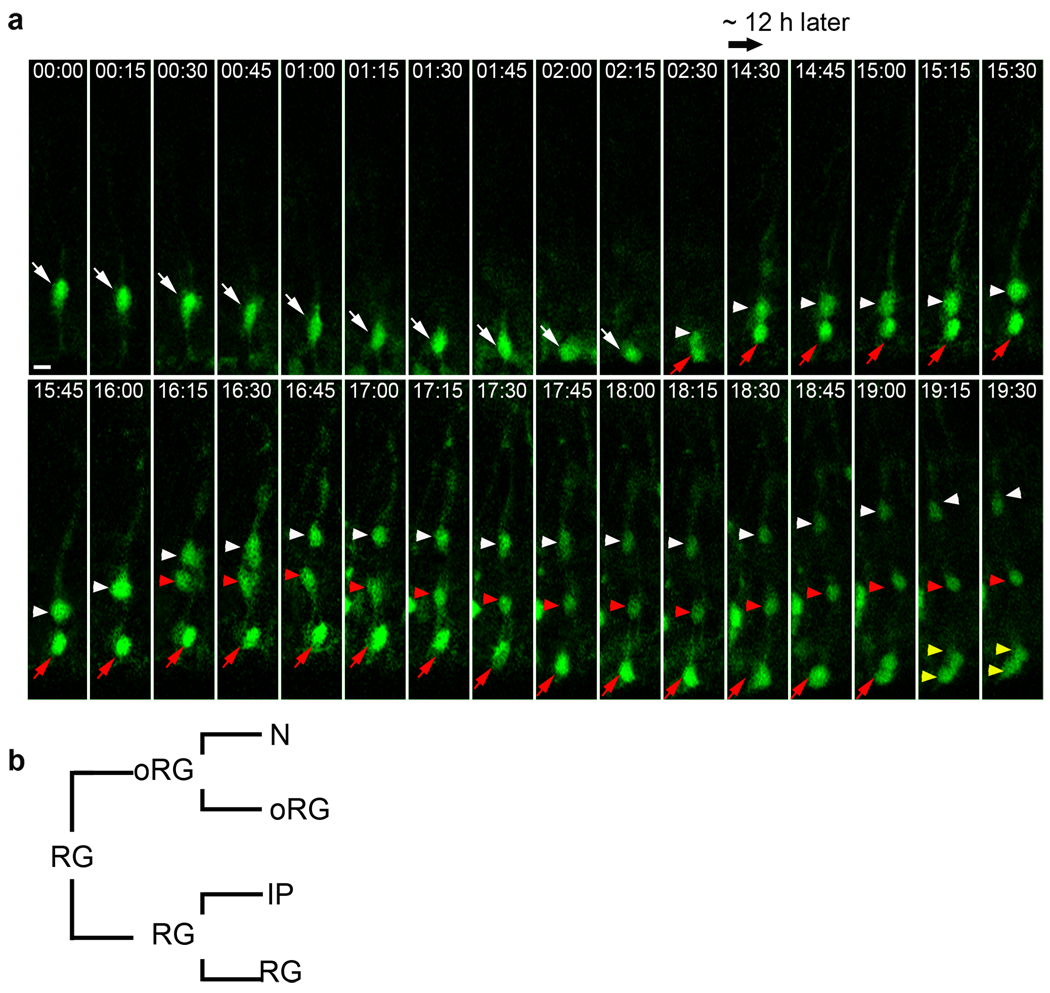
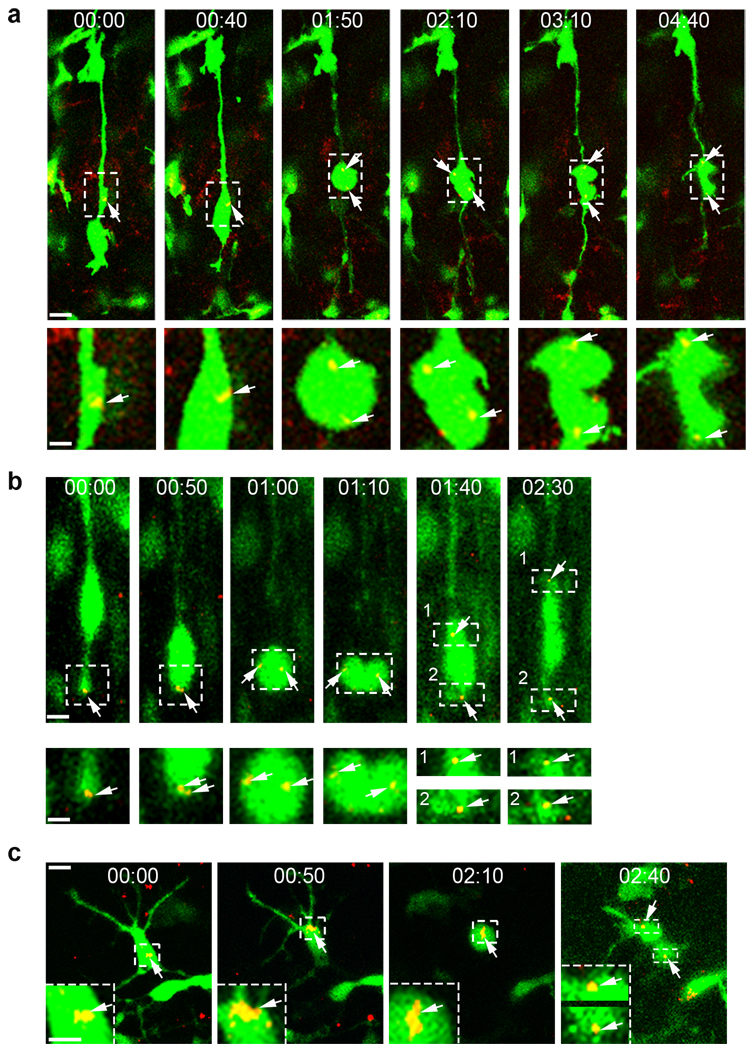
A hallmark of mammalian brain evolution is cortical expansion, which reflects an increase in the number of cortical neurons established by the progenitor cell subtypes present and the number of their neurogenic divisions. Recent studies have revealed a new class of radial glia-like (oRG) progenitor cells in the human brain, which reside in the outer subventricular zone. Expansion of the subventricular zone and appearance of oRG cells may have been essential evolutionary steps leading from lissencephalic to gyrencephalic neocortex. Here we show that oRG-like progenitor cells are present in the mouse embryonic neocortex. They arise from asymmetric divisions of radial glia and undergo self-renewing asymmetric divisions to generate neurons. Moreover, mouse oRG cells undergo mitotic somal translocation whereby centrosome movement into the basal process during interphase precedes nuclear translocation. Our finding of oRG cells in the developing rodent brain fills a gap in our understanding of neocortical expansion.
Figures
Comment in
-
Hanging by the tail: progenitor populations proliferate.Nat Neurosci. 2011 May;14(5):538-40. doi: 10.1038/nn.2817. Nat Neurosci. 2011. PMID: 21522143 No abstract available.
References
-
- Fishell G, Kriegstein AR. Neurons from radial glia: the consequences of asymmetric inheritance. Curr Opin Neurobiol. 2003;13:34–41. - PubMed
-
- Kriegstein A, Noctor S, Martinez-Cerdeno V. Patterns of neural stem and progenitor cell division may underlie evolutionary cortical expansion. Nat Rev Neurosci. 2006;7:883–890. - PubMed
-
- Rakic P. Specification of cerebral cortical areas. Science. 1988;241:170–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
