

Fusing structure and function: a structural view of the herpesvirus entry machinery
- PMID: 21478902
- PMCID: PMC3242325
- DOI: 10.1038/nrmicro2548
Fusing structure and function: a structural view of the herpesvirus entry machinery
Abstract
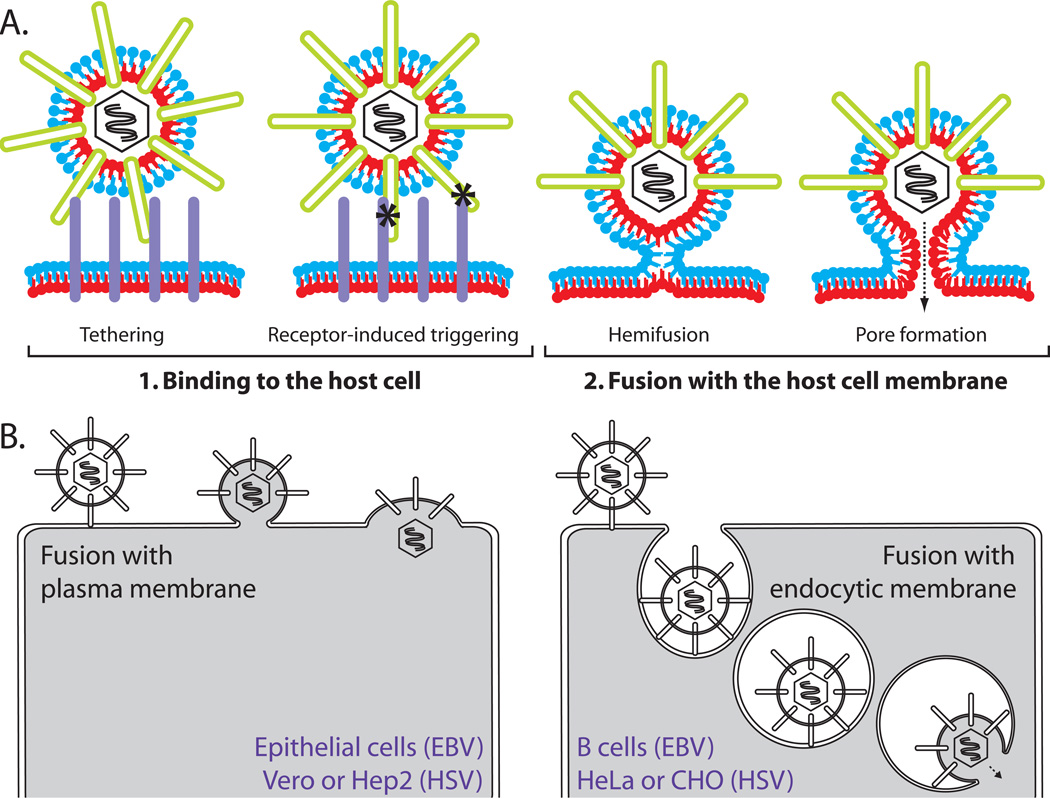
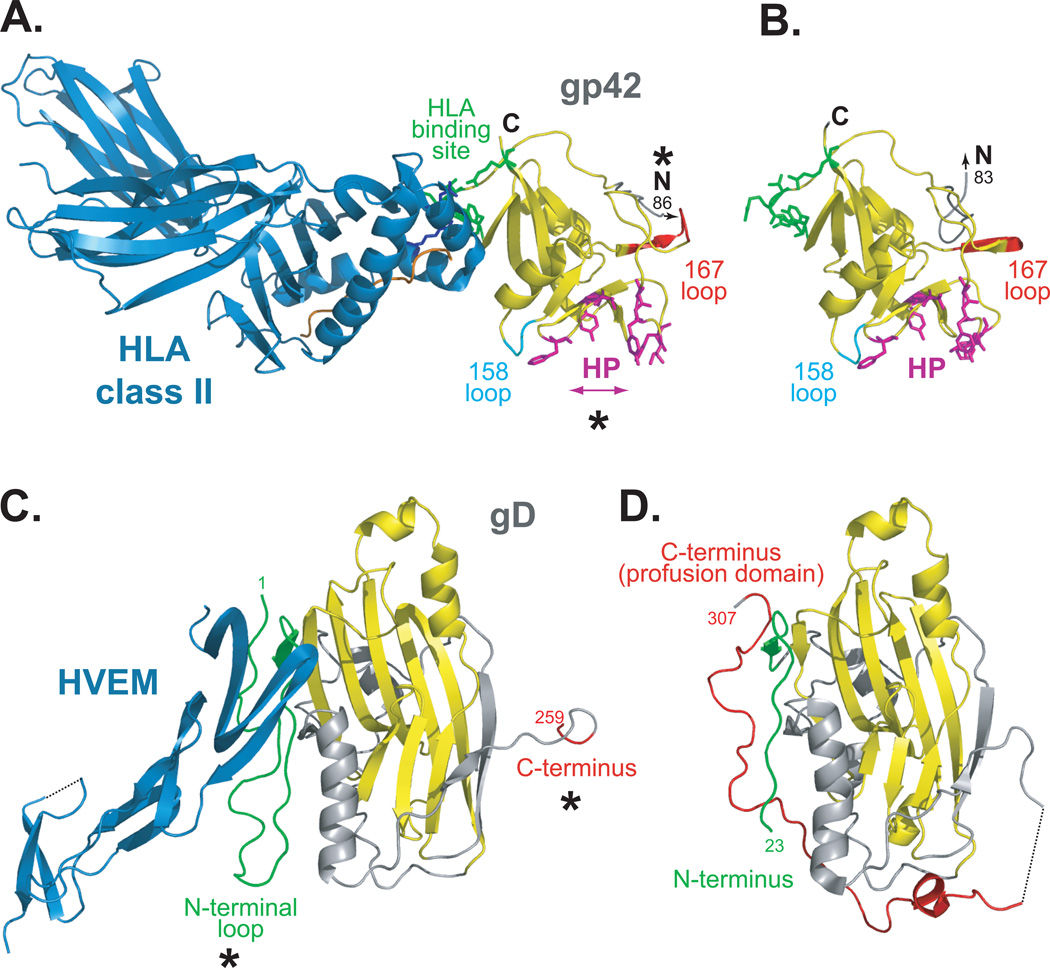
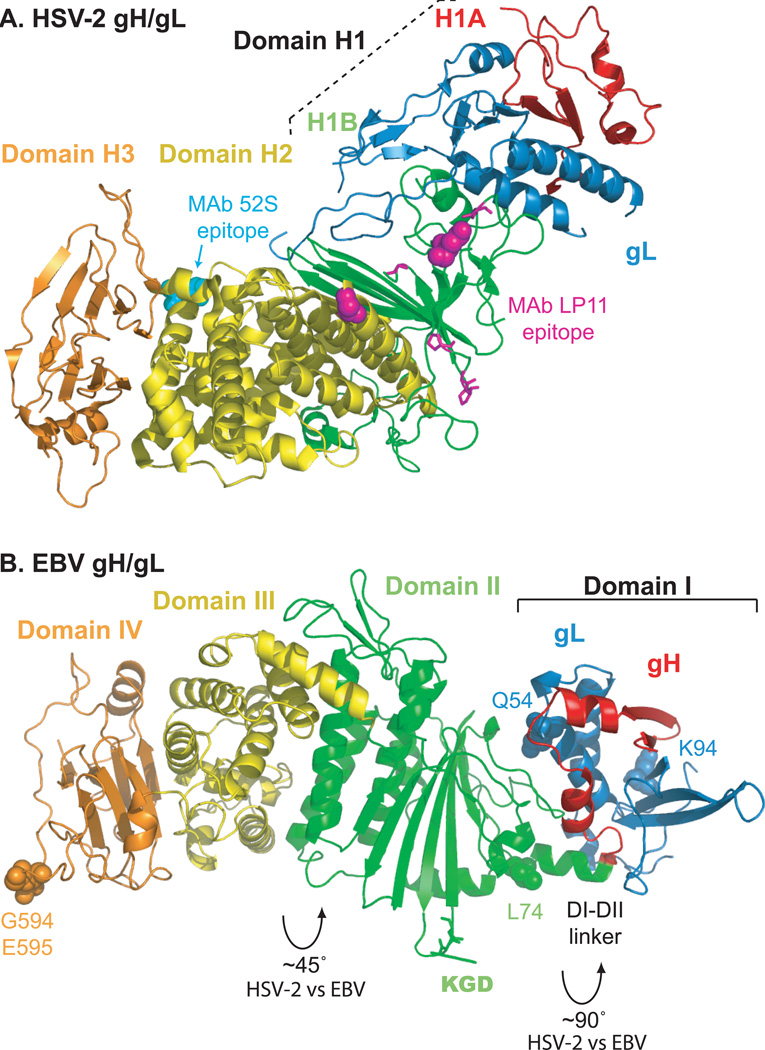
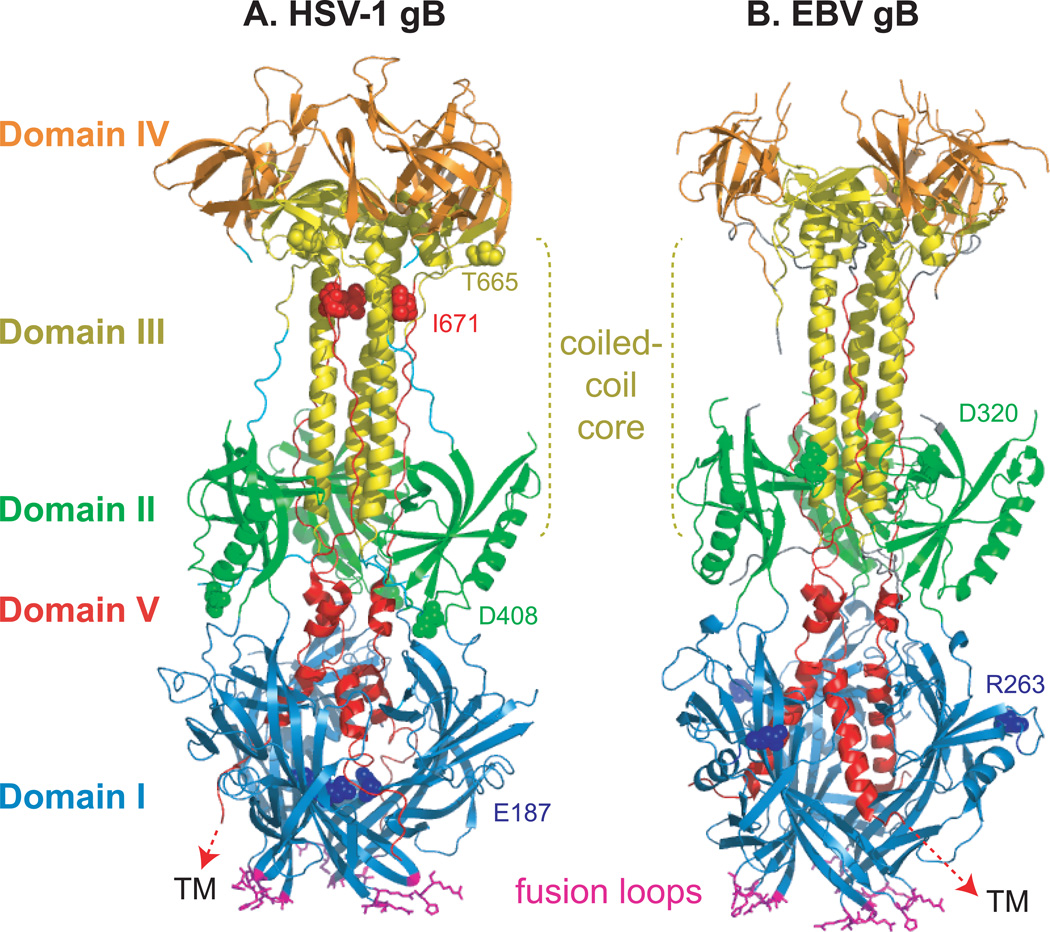
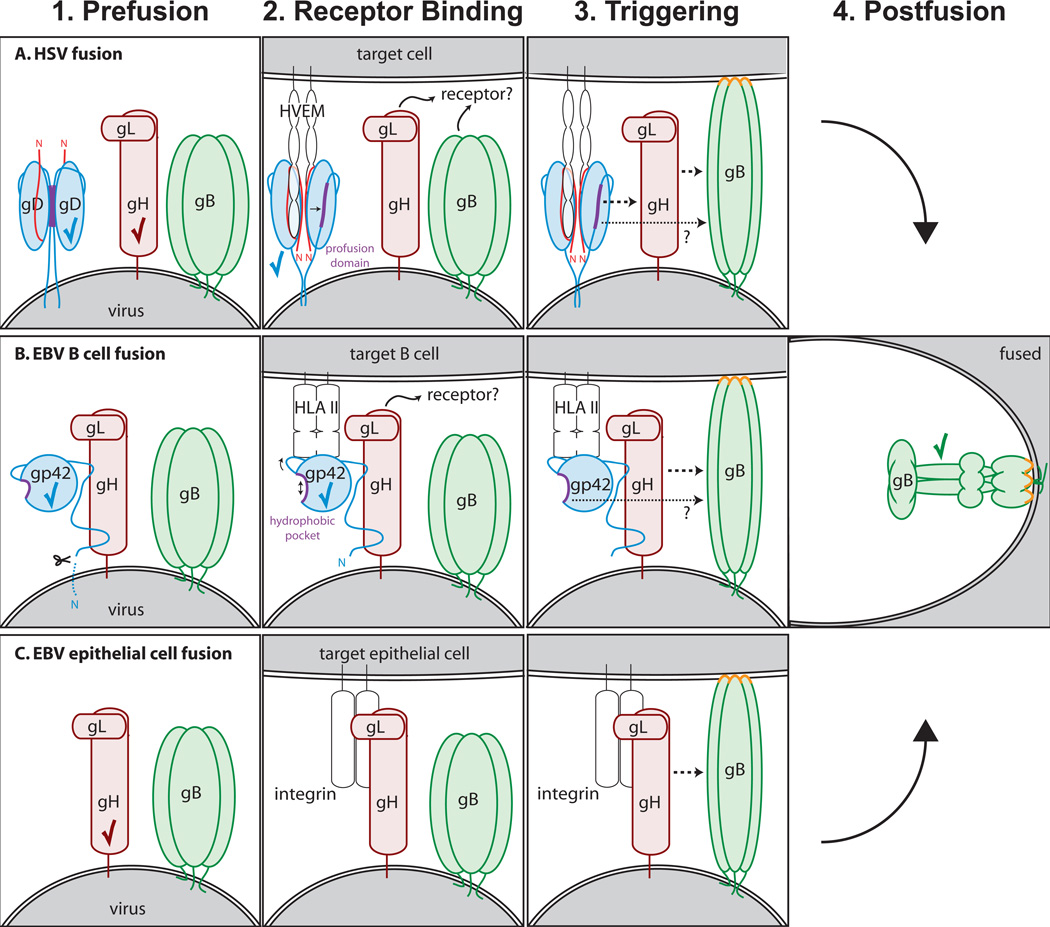
Herpesviruses are double-stranded DNA, enveloped viruses that infect host cells through fusion with either the host cell plasma membrane or endocytic vesicle membranes. Efficient infection of host cells by herpesviruses is remarkably more complex than infection by other viruses, as it requires the concerted effort of multiple glycoproteins and involves multiple host receptors. The structures of the major viral glycoproteins and a number of host receptors involved in the entry of the prototypical herpesviruses, the herpes simplex viruses (HSVs) and Epstein-Barr virus (EBV), are now known. These structural studies have accelerated our understanding of HSV and EBV binding and fusion by revealing the conformational changes that occur on virus-receptor binding, depicting potential sites of functional protein and lipid interactions, and identifying the probable viral fusogen.
Figures
References
-
- Pellet PE, Roizman B. In: Fields' Virology. 5th Edition. Knipe DM, Howley PM, editors. New York, N.Y: Lippincott-Williams and Wilkins; 2007. pp. 2479–2499.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
