

Modified needle-tip PcrV proteins reveal distinct phenotypes relevant to the control of type III secretion and intoxication by Pseudomonas aeruginosa
- PMID: 21479247
- PMCID: PMC3066235
- DOI: 10.1371/journal.pone.0018356
Modified needle-tip PcrV proteins reveal distinct phenotypes relevant to the control of type III secretion and intoxication by Pseudomonas aeruginosa
Abstract
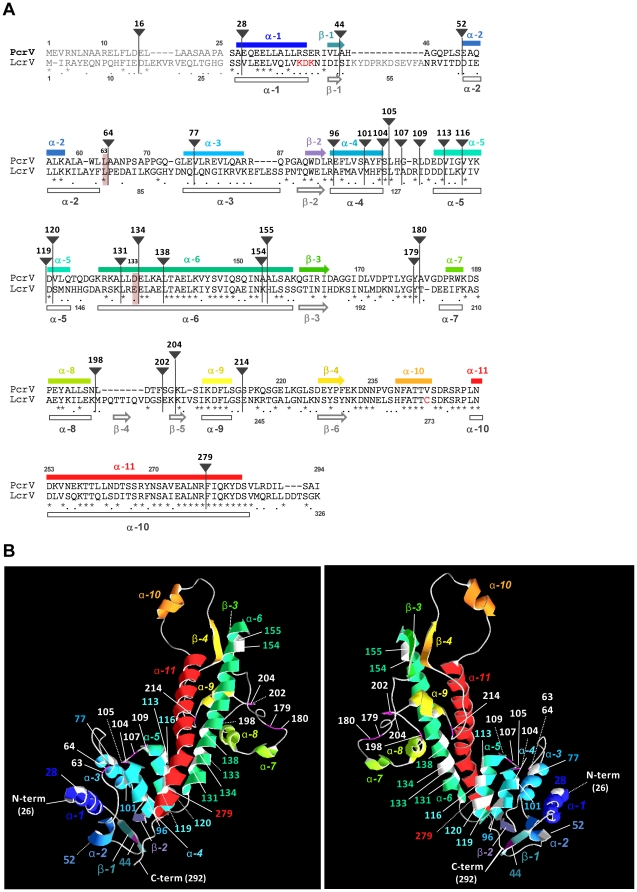
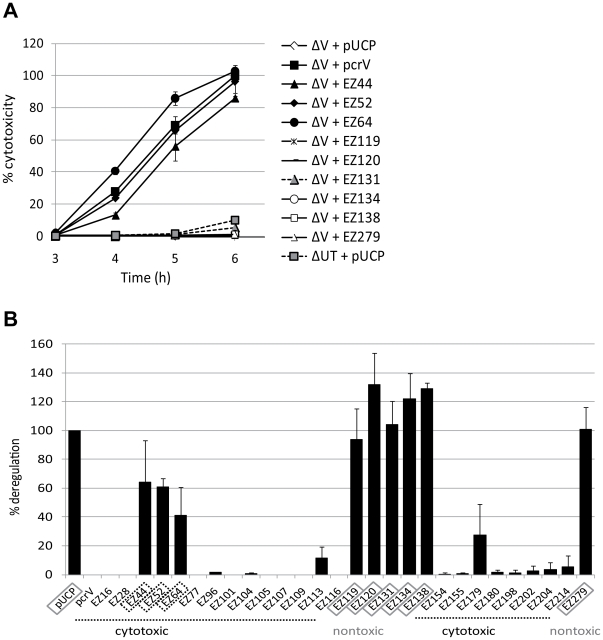
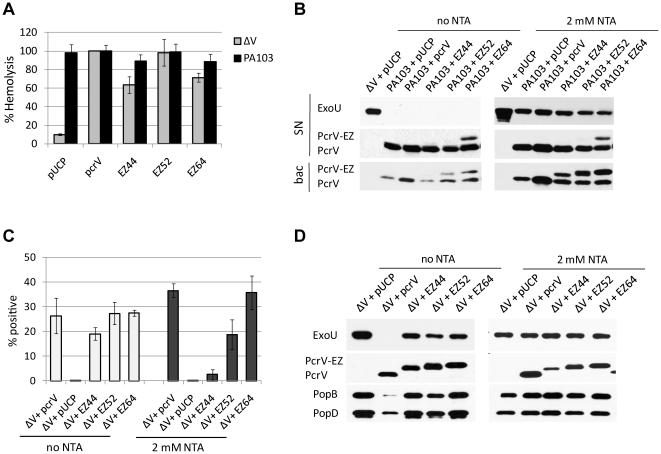
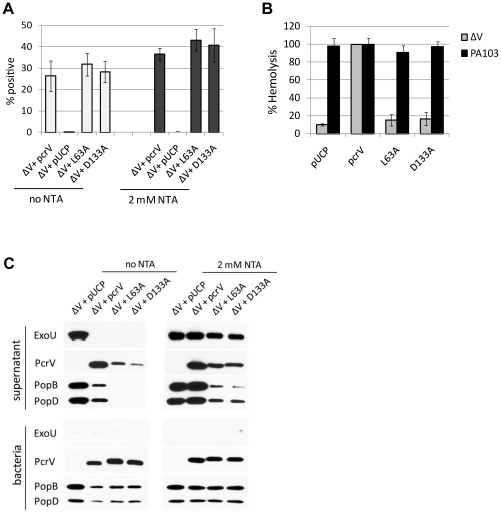
The type III secretion system (T3SS) is employed to deliver effector proteins to the cytosol of eukaryotic hosts by multiple species of Gram-negative bacteria, including Pseudomonas aeruginosa. Translocation of effectors is dependent on the proteins encoded by the pcrGVHpopBD operon. These proteins form a T3S translocator complex, composed of a needle-tip complex (PcrV), translocons (PopB and PopD), and chaperones (PcrG and PcrH). PcrV mediates the folding and insertion of PopB/PopD in host plasmic membranes, where assembled translocons form a translocation channel. Assembly of this complex and delivery of effectors through this machinery is tightly controlled by PcrV, yet the multifunctional aspects of this molecule have not been defined. In addition, PcrV is a protective antigen for P. aeruginosa infection as is the ortholog, LcrV, for Yersinia. We constructed PcrV derivatives containing in-frame linker insertions and site-specific mutations. The expression of these derivatives was regulated by a T3S-specific promoter in a pcrV-null mutant of PA103. Nine derivatives disrupted the regulation of effector secretion and constitutively released an effector protein into growth medium. Three of these regulatory mutants, in which the linker was inserted in the N-terminal globular domain, were competent for the translocation of a cytotoxin, ExoU, into eukaryotic host cells. We also isolated strains expressing a delayed-toxicity phenotype, which secrete translocators slowly despite the normal level of effector secretion. Most of the cytotoxic translocation-competent strains retained the protective epitope of PcrV derivatives, and Mab166 was able to protect erythrocytes during infection with these strains. The use of defined PcrV derivatives possessing distinct phenotypes may lead to a better understanding of the functional aspects of T3 needle-tip proteins and the development of therapeutic agents or vaccines targeting T3SS-mediated intoxication.
Conflict of interest statement
Figures




Similar articles
-
The V antigen of Pseudomonas aeruginosa is required for assembly of the functional PopB/PopD translocation pore in host cell membranes.Infect Immun. 2004 Aug;72(8):4741-50. doi: 10.1128/IAI.72.8.4741-4750.2004. Infect Immun. 2004. PMID: 15271936 Free PMC article.
-
Protein binding between PcrG-PcrV and PcrH-PopB/PopD encoded by the pcrGVH-popBD operon of the Pseudomonas aeruginosa type III secretion system.Infect Immun. 2003 Apr;71(4):2230-3. doi: 10.1128/IAI.71.4.2230-2233.2003. Infect Immun. 2003. PMID: 12654846 Free PMC article.
-
Type III secretion proteins PcrV and PcrG from Pseudomonas aeruginosa form a 1:1 complex through high affinity interactions.BMC Microbiol. 2003 Oct 18;3:21. doi: 10.1186/1471-2180-3-21. BMC Microbiol. 2003. PMID: 14565848 Free PMC article.
-
Regulation of the Yersinia type III secretion system: traffic control.Front Cell Infect Microbiol. 2013 Feb 6;3:4. doi: 10.3389/fcimb.2013.00004. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 23390616 Free PMC article. Review.
-
Structure and function of the Type III secretion system of Pseudomonas aeruginosa.Curr Protein Pept Sci. 2012 Dec;13(8):831-42. doi: 10.2174/138920312804871210. Curr Protein Pept Sci. 2012. PMID: 23305368 Free PMC article. Review.
Cited by
-
Novel humanized anti-PcrV monoclonal antibody COT-143 protects mice from lethal Pseudomonas aeruginosa infection via inhibition of toxin translocation by the type III secretion system.Antimicrob Agents Chemother. 2024 Oct 8;68(10):e0069424. doi: 10.1128/aac.00694-24. Epub 2024 Sep 13. Antimicrob Agents Chemother. 2024. PMID: 39269189 Free PMC article.
-
Identification of broadly protective human antibodies to Pseudomonas aeruginosa exopolysaccharide Psl by phenotypic screening.J Exp Med. 2012 Jul 2;209(7):1273-87. doi: 10.1084/jem.20120033. Epub 2012 Jun 25. J Exp Med. 2012. PMID: 22734046 Free PMC article.
-
Intoxication of host cells by the T3SS phospholipase ExoU: PI(4,5)P2-associated, cytoskeletal collapse and late phase membrane blebbing.PLoS One. 2014 Jul 25;9(7):e103127. doi: 10.1371/journal.pone.0103127. eCollection 2014. PLoS One. 2014. PMID: 25061861 Free PMC article.
-
Surface organelles assembled by secretion systems of Gram-negative bacteria: diversity in structure and function.FEMS Microbiol Rev. 2012 Nov;36(6):1046-82. doi: 10.1111/j.1574-6976.2012.00342.x. Epub 2012 May 24. FEMS Microbiol Rev. 2012. PMID: 22545799 Free PMC article. Review.
-
In the absence of effector proteins, the Pseudomonas aeruginosa type three secretion system needle tip complex contributes to lung injury and systemic inflammatory responses.PLoS One. 2013 Nov 27;8(11):e81792. doi: 10.1371/journal.pone.0081792. eCollection 2013. PLoS One. 2013. PMID: 24312357 Free PMC article.
References
-
- Giamarellou H. Therapeutic guidelines for Pseudomonas aeruginosa infections. Int J Antimicrob Agents. 2000;16:103–106. - PubMed
-
- Lyczak JB, Cannon CL, Pier GB. Establishment of Pseudomonas aeruginosa infection: lessons from a versatile opportunist. Microbes Infect. 2000;2:1051–1060. - PubMed
-
- Crouch BS, Wunderink RG, Jones CB, Leeper KV., Jr Ventilator-associated pneumonia due to Pseudomonas aeruginosa. Chest. 1996;109:1019–1029. - PubMed
-
- Vidal F, et al. Epidemiology and outcome of Pseudomonas aeruginosa bacteremia, with special emphasis on the influence of antibiotic treatment. Analysis of 189 episodes. Arch Intern Med. 1996;156:2121–2126. - PubMed
-
- Roy-Burman A, et al. Type III protein secretion is associated with death in lower respiratory and systemic Pseudomonas aeruginosa infections. J Infect Dis. 2001;183:1767–1774. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
