

Rap1a activation by CalDAG-GEFI and p38 MAPK is involved in E-selectin-dependent slow leukocyte rolling
- PMID: 21480213
- PMCID: PMC3124568
- DOI: 10.1002/eji.201041196
Rap1a activation by CalDAG-GEFI and p38 MAPK is involved in E-selectin-dependent slow leukocyte rolling
Abstract
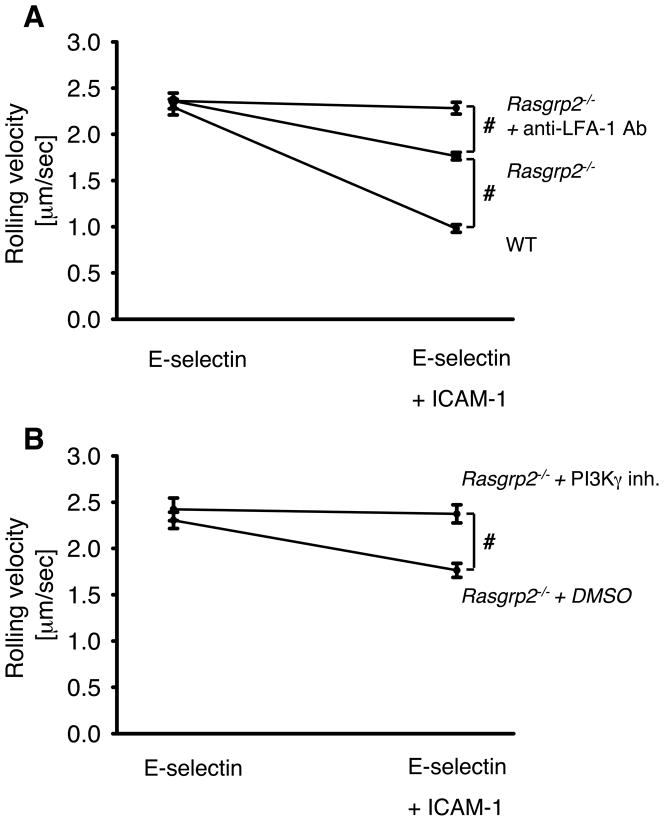
Rolling leukocytes are exposed to different adhesion molecules and chemokines. Neutrophils rolling on E-selectin induce integrin αLβ2-mediated slow rolling on ICAM-1 by activating a phospholipase C (PLC)γ2-dependent and a separate PI3Kγ-dependent pathway. E-selectin-signaling cooperates with chemokine signaling to recruit neutrophils into inflamed tissues. However, the distal signaling pathway linking PLCγ2 (Plcg2) to αLβ2-activation is unknown. To identify this pathway, we used different Tat-fusion-mutants and gene-deficient mice in intravital microscopy, autoperfused flow chamber, peritonitis, and biochemical studies. We found that the small GTPase Rap1 is activated following E-selectin engagement and that blocking Rap1a in Pik3cg-/- mice by a dominant-negative Tat-fusion mutant completely abolished E-selectin-mediated slow rolling. We identified CalDAG-GEFI (Rasgrp2) and p38 MAPK as key signaling intermediates between PLCγ2 and Rap1a. Gαi-independent leukocyte adhesion to and transmigration through endothelial cells in inflamed postcapillary venules of the cremaster muscle were completely abolished in Rasgrp2-/- mice. The physiological importance of CalDAG-GEFI in E-selectin-dependent integrin activation is shown by complete inhibition of neutrophil recruitment into the inflamed peritoneal cavity of Rasgrp2-/- leukocytes treated with pertussis toxin to block Gαi-signaling. Our data demonstrate that Rap1a activation by p38 MAPK and CalDAG-GEFI is involved in E-selectin-dependent slow rolling and leukocyte recruitment.
Copyright © 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





Similar articles
-
Tyrosine kinase Btk regulates E-selectin-mediated integrin activation and neutrophil recruitment by controlling phospholipase C (PLC) gamma2 and PI3Kgamma pathways.Blood. 2010 Apr 15;115(15):3118-27. doi: 10.1182/blood-2009-11-254185. Epub 2010 Feb 18. Blood. 2010. PMID: 20167705 Free PMC article.
-
E-selectin engages PSGL-1 and CD44 through a common signaling pathway to induce integrin alphaLbeta2-mediated slow leukocyte rolling.Blood. 2010 Jul 22;116(3):485-94. doi: 10.1182/blood-2009-12-259556. Epub 2010 Mar 18. Blood. 2010. PMID: 20299514 Free PMC article.
-
PSGL-1 engagement by E-selectin signals through Src kinase Fgr and ITAM adapters DAP12 and FcR gamma to induce slow leukocyte rolling.J Exp Med. 2008 Sep 29;205(10):2339-47. doi: 10.1084/jem.20072660. Epub 2008 Sep 15. J Exp Med. 2008. PMID: 18794338 Free PMC article.
-
Reprint of Neutrophil cell surface receptors and their intracellular signal transduction pathways.Int Immunopharmacol. 2013 Dec;17(4):1185-97. doi: 10.1016/j.intimp.2013.11.010. Epub 2013 Nov 18. Int Immunopharmacol. 2013. PMID: 24263067 Review.
-
Neutrophil cell surface receptors and their intracellular signal transduction pathways.Int Immunopharmacol. 2013 Nov;17(3):638-50. doi: 10.1016/j.intimp.2013.06.034. Epub 2013 Aug 30. Int Immunopharmacol. 2013. PMID: 23994464 Free PMC article. Review.
Cited by
-
Cardiopulmonary bypass during cardiac surgery modulates systemic inflammation by affecting different steps of the leukocyte recruitment cascade.PLoS One. 2012;7(9):e45738. doi: 10.1371/journal.pone.0045738. Epub 2012 Sep 19. PLoS One. 2012. PMID: 23029213 Free PMC article.
-
Blocking neutrophil integrin activation prevents ischemia-reperfusion injury.J Exp Med. 2015 Jul 27;212(8):1267-81. doi: 10.1084/jem.20142358. Epub 2015 Jul 13. J Exp Med. 2015. PMID: 26169939 Free PMC article.
-
Lipid Raft is required for PSGL-1 ligation induced HL-60 cell adhesion on ICAM-1.PLoS One. 2013 Dec 3;8(12):e81807. doi: 10.1371/journal.pone.0081807. eCollection 2013. PLoS One. 2013. PMID: 24312591 Free PMC article.
-
Leukocyte integrins: role in leukocyte recruitment and as therapeutic targets in inflammatory disease.Pharmacol Ther. 2015 Mar;147:123-135. doi: 10.1016/j.pharmthera.2014.11.008. Epub 2014 Nov 14. Pharmacol Ther. 2015. PMID: 25448040 Free PMC article. Review.
-
Extracellular MRP8/14 is a regulator of β2 integrin-dependent neutrophil slow rolling and adhesion.Nat Commun. 2015 Apr 20;6:6915. doi: 10.1038/ncomms7915. Nat Commun. 2015. PMID: 25892652 Free PMC article.
References
-
- Anderson DC, Springer TA. Leukocyte adhesion deficiency: an inherited defect in the Mac-1, LFA-1, and p150,95 glycoproteins. Annu Rev Med. 1987;38:175–194. - PubMed
-
- Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol. 2006;6:173–182. - PubMed
-
- Butcher EC. Leukocyte-endothelial cell recognition: three (or more) steps to specificity and diversity. Cell. 1991;67:1033–1036. - PubMed
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76:301–314. - PubMed
-
- Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7:678–689. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
