

Traces of Drosophila memory
- PMID: 21482352
- PMCID: PMC3374581
- DOI: 10.1016/j.neuron.2011.03.012
Traces of Drosophila memory
Abstract
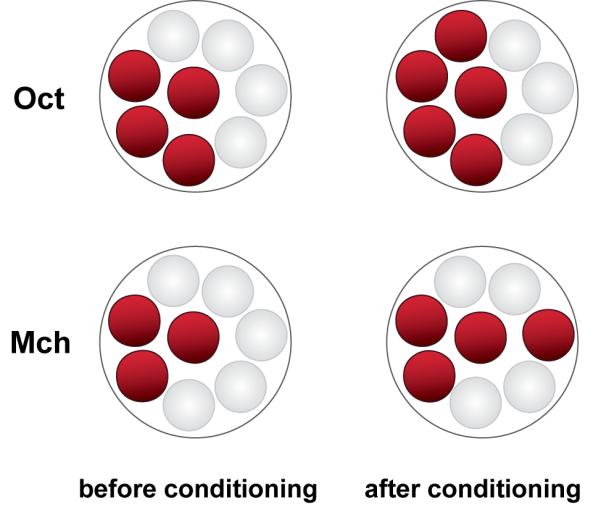
Studies using functional cellular imaging of living flies have identified six memory traces that form in the olfactory nervous system after conditioning with odors. These traces occur in distinct nodes of the olfactory nervous system, form and disappear across different windows of time, and are detected in the imaged neurons as increased calcium influx or synaptic release in response to the conditioned odor. Three traces form at or near acquisition and coexist with short-term behavioral memory. One trace forms with a delay after learning and coexists with intermediate-term behavioral memory. Two traces form many hours after acquisition and coexist with long-term behavioral memory. The transient memory traces may support behavior across the time windows of their existence. The experimental approaches for dissecting memory formation in the fly, ranging from the molecular to the systems, make it an ideal system for elucidating the logic by which the nervous system organizes and stores different temporal forms of memory.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures








References
-
- Ashraf SI, McLoon AL, Sclarsic SM, Kunes S. Synaptic protein synthesis associated with memory is regulated by the RISC pathway in Drosophlia. Cell. 2006;124:191–205. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
