

Epigenetic mechanisms in Alzheimer's disease
- PMID: 21482442
- PMCID: PMC3115415
- DOI: 10.1016/j.neurobiolaging.2010.08.017
Epigenetic mechanisms in Alzheimer's disease
Abstract
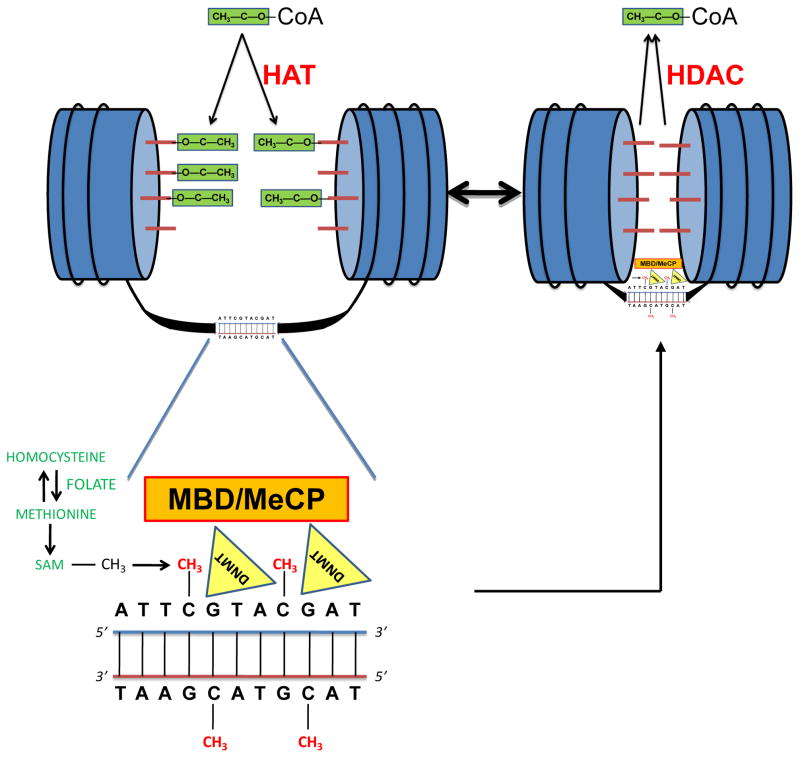
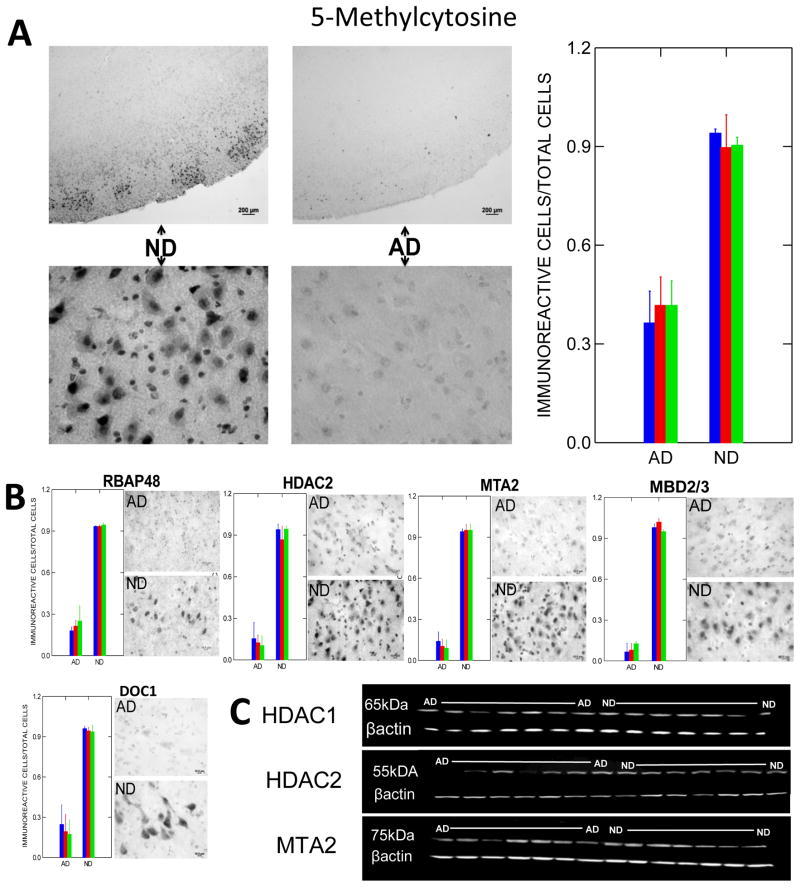
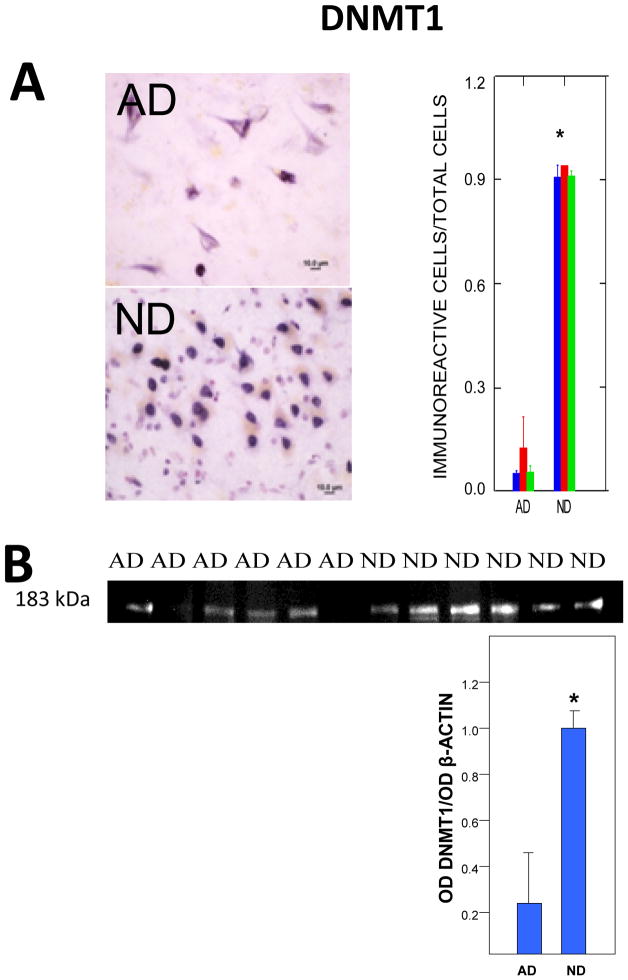
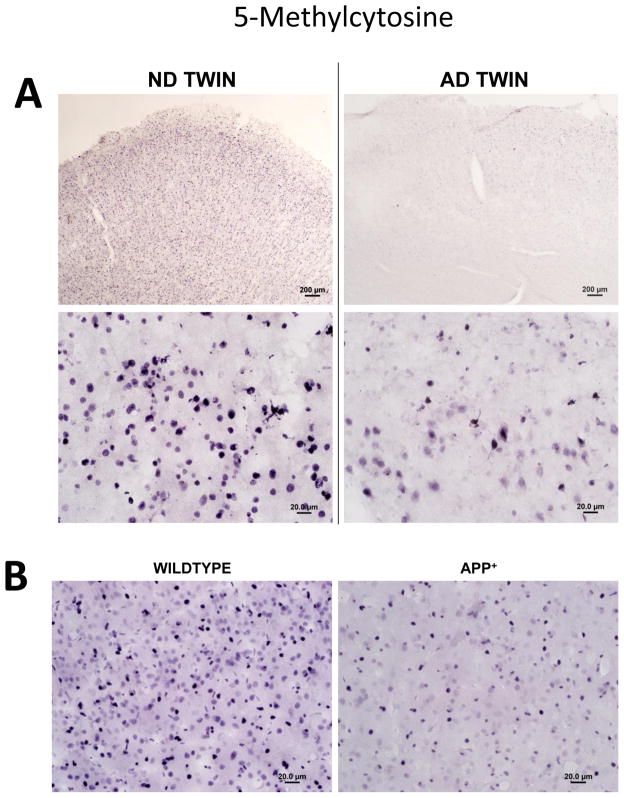
Epigenetic modifications help orchestrate sweeping developmental, aging, and disease-causing changes in phenotype by altering transcriptional activity in multiple genes spanning multiple biologic pathways. Although previous epigenetic research has focused primarily on dividing cells, particularly in cancer, recent studies have shown rapid, dynamic, and persistent epigenetic modifications in neurons that have significant neuroendocrine, neurophysiologic, and neurodegenerative consequences. Here, we provide a review of the major mechanisms for epigenetic modification and how they are reportedly altered in aging and Alzheimer's disease (AD). Because of their reach across the genome, epigenetic mechanisms may provide a unique integrative framework for the pathologic diversity and complexity of AD.
Copyright © 2011 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors state that they have no actual or potential conflict of interest that could inappropriately influence this work.
Figures
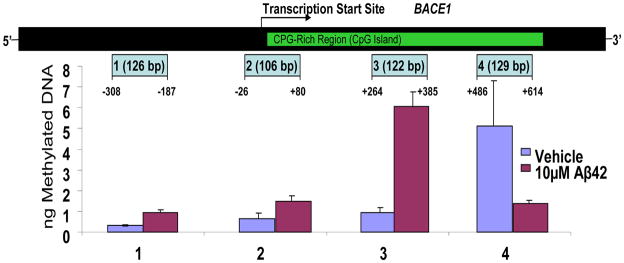
Comment in
-
Toward an integrated genetic and epigenetic approach to Alzheimer's disease.Neurobiol Aging. 2011 Jul;32(7):1188-91. doi: 10.1016/j.neurobiolaging.2010.10.021. Epub 2011 Apr 16. Neurobiol Aging. 2011. PMID: 21497948 Review.
-
One-carbon metabolism and Alzheimer's disease: is it all a methylation matter?Neurobiol Aging. 2011 Jul;32(7):1192-5. doi: 10.1016/j.neurobiolaging.2011.01.012. Epub 2011 Apr 23. Neurobiol Aging. 2011. PMID: 21524430 Review.
-
Epigenetic mechanisms in Alzheimer's disease: progress but much to do.Neurobiol Aging. 2011 Jul;32(7):1181-7. doi: 10.1016/j.neurobiolaging.2011.02.024. Neurobiol Aging. 2011. PMID: 21669333 Review.
References
-
- Abelson JF, Kwan KY, O’Roak BJ, Baek DY, Stillman AA, Morgan TM, Mathews CA, Pauls DL, Rasin M-R, Gunel M, Spertus JA, Leckman JF, Dure LS, Kurlan R, Singer HS, Gilbert DL, Farhi A, Louvi A, Lifton RP, Sestan N, State MW. Sequence variants in SLITRK1 are associated with Tourette’s syndrome. Science. 2005;310:317–320. - PubMed
-
- Agrawal A, Murphy RF, Agrawal DK. DNA methylation in breast and colorectal cancers. Mod Pathol. 2007;20:711–721. - PubMed
-
- Aizawa S, Yamamuro Y. Involvement of histone acetylation in the regulation of choline acetyltransferase gene in NG108-15 neuronal cells. Neurochem Int. 2010 [Epub ahead of print] - PubMed
-
- Allfrey VG. Structural modifications of histones and their possible role in the regulation of ribonucleic acid synthesis. Proc Can Cancer Conf. 1966;6:313–335. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
