

Mediator influences telomeric silencing and cellular life span
- PMID: 21482672
- PMCID: PMC3133415
- DOI: 10.1128/MCB.05242-11
Mediator influences telomeric silencing and cellular life span
Abstract
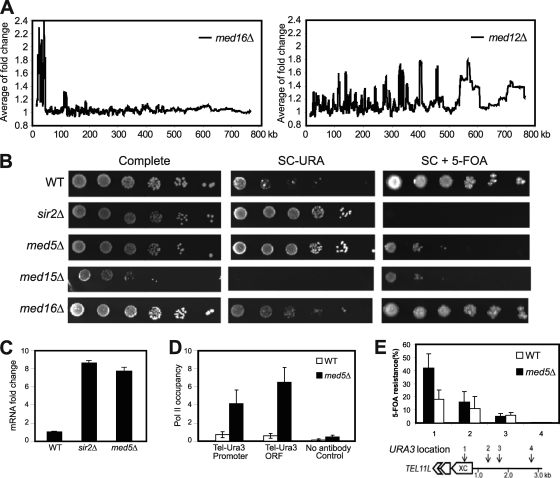
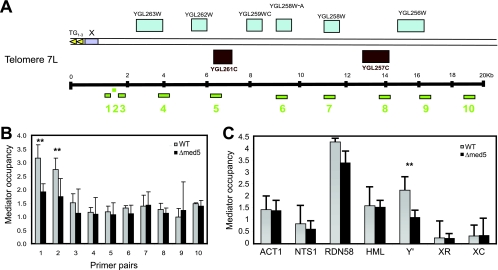
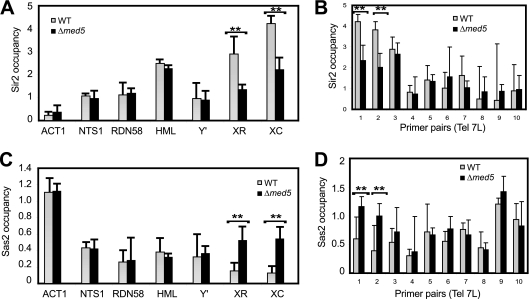
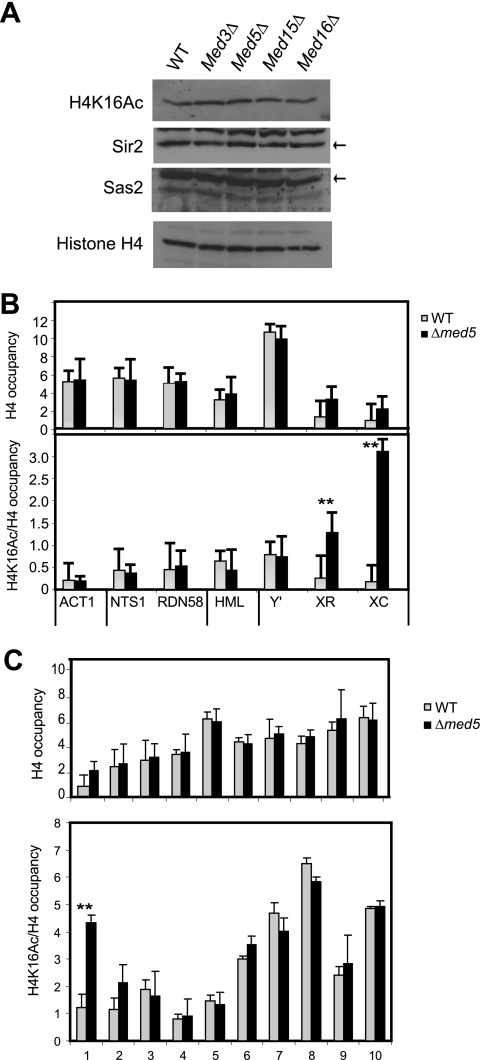
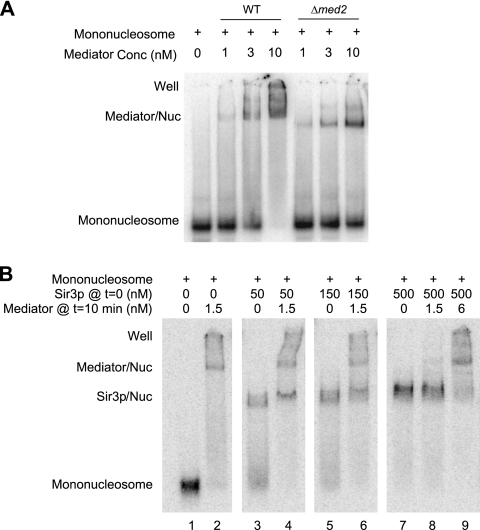
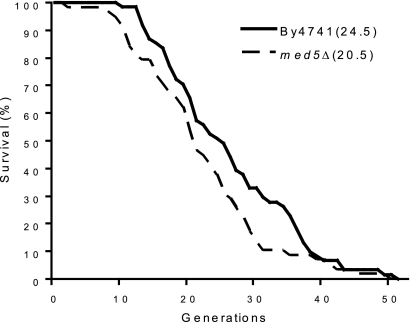
The Mediator complex is required for the regulated transcription of nearly all RNA polymerase II-dependent genes. Here we demonstrate a new role for Mediator which appears to be separate from its function as a transcriptional coactivator. Mediator associates directly with heterochromatin at telomeres and influences the exact boundary between active and inactive chromatin. Loss of the Mediator Med5 subunit or mutations in Med7 cause a depletion of the complex from regions located near subtelomeric X elements, which leads to a change in the balance between the Sir2 and Sas2 proteins. These changes in turn result in increased levels of H4K16 acetylation near telomeres and in desilencing of subtelomeric genes. Increases in H4K16 acetylation have been observed at telomeres in aging cells. In agreement with this observation, we found that the loss of MED5 leads to shortening of the Saccharomyces cerevisiae (budding yeast) replicative life span.
Figures
References
-
- Andrau J. C., et al. 2006. Genome-wide location of the coactivator mediator: binding without activation and transient Cdk8 interaction on DNA. Mol. Cell 22: 179–192 - PubMed
-
- Baidoobonso S. M., Guidi B. W., Myers L. C. 2007. Med19(Rox3) regulates intermodule interactions in the Saccharomyces cerevisiae mediator complex. J. Biol. Chem. 282: 5551–5559 - PubMed
-
- Beve J., et al. 2005. The structural and functional role of Med5 in the yeast Mediator tail module. J. Biol. Chem. 280: 41366–41372 - PubMed
-
- Bjorklund S., Gustafsson C. M. 2004. The mediator complex. Adv. Protein Chem. 67: 43–65 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases