

Overexpression of cdk4 and cyclinD1 triggers greater expansion of neural stem cells in the adult mouse brain
- PMID: 21482697
- PMCID: PMC3092341
- DOI: 10.1084/jem.20102167
Overexpression of cdk4 and cyclinD1 triggers greater expansion of neural stem cells in the adult mouse brain
Abstract
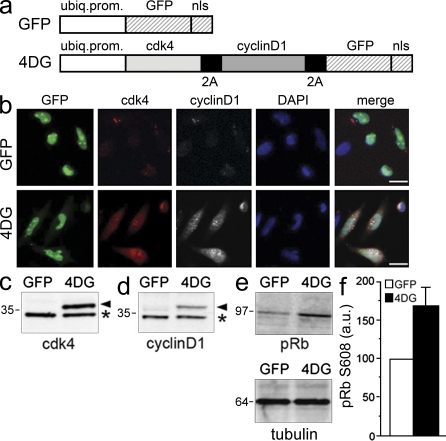
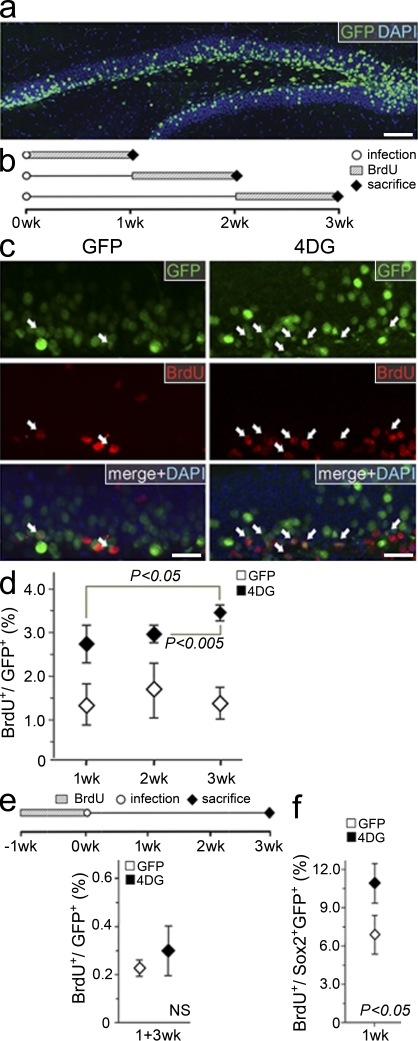
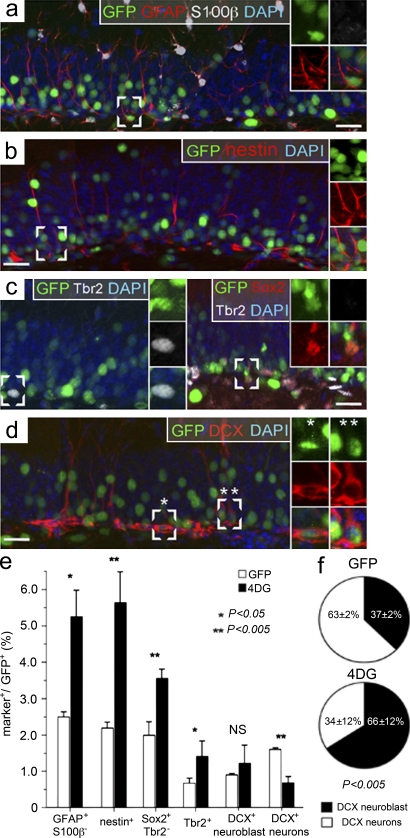
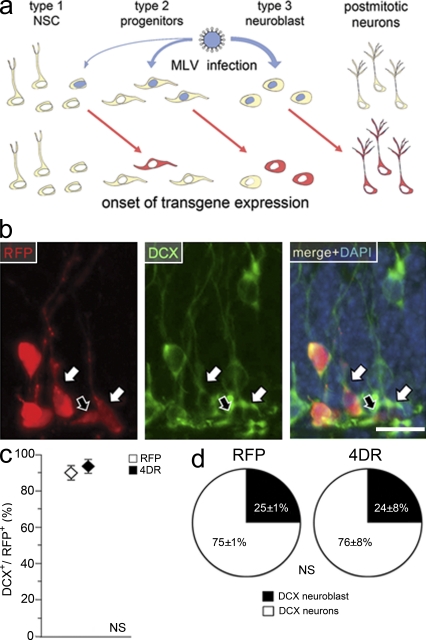
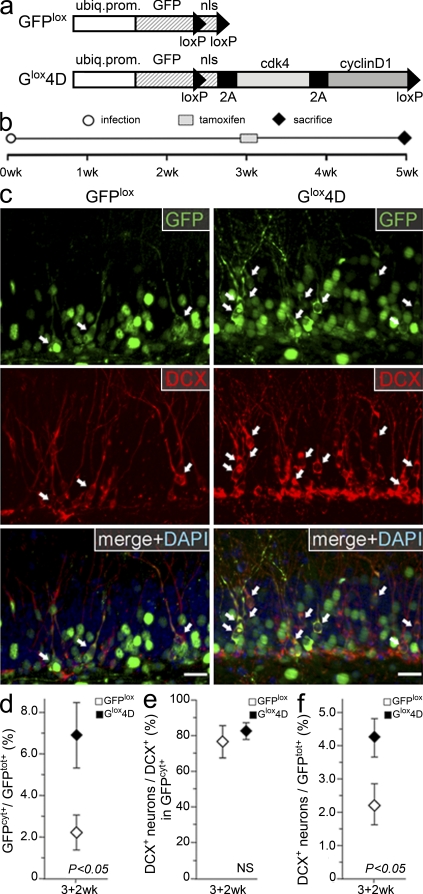
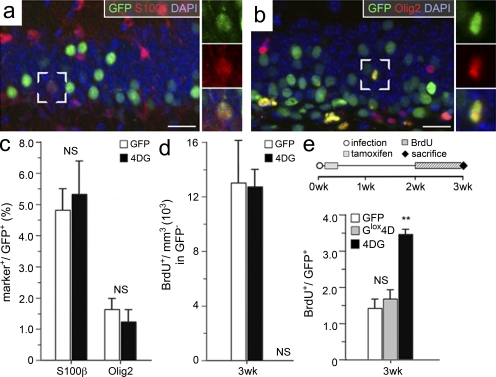
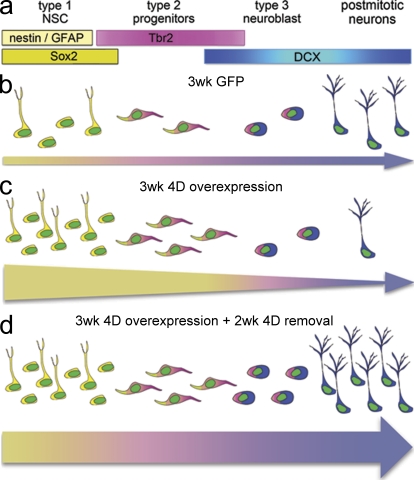
Neural stem cells (NSCs) in the adult mammalian brain generate neurons and glia throughout life. However, the physiological role of adult neurogenesis and the use of NSCs for therapy are highly controversial. One factor hampering the study and manipulation of neurogenesis is that NSCs, like most adult somatic stem cells, are difficult to expand and their switch to differentiation is hard to control. In this study, we show that acute overexpression of the cdk4 (cyclin-dependent kinase 4)-cyclinD1 complex in the adult mouse hippocampus cell-autonomously increases the expansion of neural stem and progenitor cells while inhibiting neurogenesis. Importantly, we developed a system that allows the temporal control of cdk4-cyclinD1 overexpression, which can be used to increase the number of neurons generated from the pool of manipulated precursor cells. Beside providing a proof of principle that expansion versus differentiation of somatic stem cells can be controlled in vivo, our study describes, to the best of our knowledge, the first acute and inducible temporal control of neurogenesis in the mammalian brain, which may be critical for identifying the role of adult neurogenesis, using NSCs for therapy, and, perhaps, extending our findings to other adult somatic stem cells.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
